S-ar putea să vă placă și
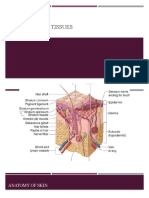
- Anatomy and Physiology of The Skin.3Document11 paginiAnatomy and Physiology of The Skin.3alinaÎncă nu există evaluări
- 1 SS Skin Cancer - Chapter 1Document12 pagini1 SS Skin Cancer - Chapter 1Muhammad IqbalÎncă nu există evaluări
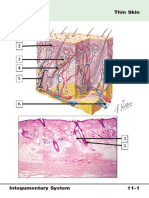
- Anatomy and Physiology of The SkinDocument30 paginiAnatomy and Physiology of The SkinNancy VargasÎncă nu există evaluări
- Anatomy and Physiology of SkinDocument12 paginiAnatomy and Physiology of Skinjose abadÎncă nu există evaluări
- Bab 7. Struktur KulitDocument56 paginiBab 7. Struktur KulityumizoneÎncă nu există evaluări
- Core Curriculum WOCNSDocument1.212 paginiCore Curriculum WOCNSJerry MaguireÎncă nu există evaluări
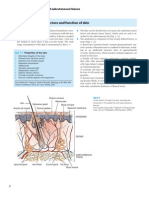
- Anatomy and Physiology: Key FeaturesDocument4 paginiAnatomy and Physiology: Key Featuresorchitarosadila haryonoÎncă nu există evaluări
- ANA 101 Lab: University of Northern PhilippinesDocument12 paginiANA 101 Lab: University of Northern PhilippinesAesthethic findsÎncă nu există evaluări
- Week 4 Case StudyDocument6 paginiWeek 4 Case StudyElizabeth GithinjiÎncă nu există evaluări
- Vertebrate Integument and DerivativesDocument16 paginiVertebrate Integument and DerivativesNarasimha MurthyÎncă nu există evaluări
- BM1011 2020 Study Guide For Module 3 The Integumentary SystemDocument14 paginiBM1011 2020 Study Guide For Module 3 The Integumentary SystemLaetitia ReyesÎncă nu există evaluări
- GENERAL HISTOLOGY pdf-1Document22 paginiGENERAL HISTOLOGY pdf-1Dany Samuel100% (1)
- 1.integumentary System - HistologyDocument34 pagini1.integumentary System - Histologyqty9jgkpnzÎncă nu există evaluări
- Lab Activity - DMSDocument85 paginiLab Activity - DMSPRAMESTI WARDHANIÎncă nu există evaluări
- Skin LayersDocument3 paginiSkin LayersCrackmcat CrackmcatÎncă nu există evaluări
- Integumentary System: Stratum BasaleDocument21 paginiIntegumentary System: Stratum BasalerinaÎncă nu există evaluări
- Characteristics of Living Organisms and The Integumentary SystemDocument11 paginiCharacteristics of Living Organisms and The Integumentary SystemSHYRE COLEEN CATAYAOÎncă nu există evaluări
- Skin Structure and FunctionDocument4 paginiSkin Structure and FunctionPutri ClaraÎncă nu există evaluări
- 02 Kandarova03Document31 pagini02 Kandarova03ضحى الغزيÎncă nu există evaluări
- Anatomy, Skin (Integument), Epidermis - StatPearls - NCBI BookshelfDocument8 paginiAnatomy, Skin (Integument), Epidermis - StatPearls - NCBI Bookshelfpka25Încă nu există evaluări
- Anatomy Histo Physio DR - AdilDocument73 paginiAnatomy Histo Physio DR - AdilHybat ElsheikhÎncă nu există evaluări
- Anatomy and PhysiologyDocument25 paginiAnatomy and PhysiologyNaidin Catherine De Guzman-AlcalaÎncă nu există evaluări
- Topical Corticosteroids BinderDocument108 paginiTopical Corticosteroids BinderElena DragomirÎncă nu există evaluări
- IntegumentaryDocument7 paginiIntegumentarySimone ApostolÎncă nu există evaluări
- Basic Physiology of The SkinDocument4 paginiBasic Physiology of The SkinRegina SinagaÎncă nu există evaluări
- Technological Institute of The PhilippinesDocument8 paginiTechnological Institute of The PhilippinesJohn Dolph FacundoÎncă nu există evaluări
- Anatomy, Skin (Integument) - StatPearls - NCBI BookshelfDocument4 paginiAnatomy, Skin (Integument) - StatPearls - NCBI Bookshelfputri aisheÎncă nu există evaluări
- Epidermis Dermis Hypodermis or Subcutaneous FatDocument4 paginiEpidermis Dermis Hypodermis or Subcutaneous FatAyesha RathÎncă nu există evaluări
- Skin Structure and Function: Figure 1.1 Cross-Section of The SkinDocument11 paginiSkin Structure and Function: Figure 1.1 Cross-Section of The SkinfunmugÎncă nu există evaluări
- Laporan Pendahuluan Asuhan Keperawatan Skin Loss A. PengertianDocument9 paginiLaporan Pendahuluan Asuhan Keperawatan Skin Loss A. PengertianPrasetyo official.01Încă nu există evaluări
- The Potential of Metabolic Interventions To Enhance Transdermal Drug DeliveryDocument7 paginiThe Potential of Metabolic Interventions To Enhance Transdermal Drug DeliveryoktaviacxÎncă nu există evaluări
- Dermatology Questions and AnsDocument147 paginiDermatology Questions and AnsEl FaroukÎncă nu există evaluări
- Link To Publication in University of Groningen/UMCG Research DatabaseDocument49 paginiLink To Publication in University of Groningen/UMCG Research DatabasescribdangeloÎncă nu există evaluări
- Therapeutic Potential of Stem Cells in Skin Repair and RegenerationDocument13 paginiTherapeutic Potential of Stem Cells in Skin Repair and RegenerationTaro RahmatiaÎncă nu există evaluări
- ArchanaDocument10 paginiArchanaUthin Latif MslÎncă nu există evaluări
- Unit 5 Skin and Integumentary SystemDocument104 paginiUnit 5 Skin and Integumentary SystemSino McakoÎncă nu există evaluări
- 2017 - Tecnología CosméticaDocument54 pagini2017 - Tecnología CosméticaCésar Rosado Avila ToñoÎncă nu există evaluări
- ANA 222 Intergumentry System HistologyDocument7 paginiANA 222 Intergumentry System HistologyprincessmakklisÎncă nu există evaluări
- The Integumentary SystemDocument6 paginiThe Integumentary SystemWolverineInZenÎncă nu există evaluări
- Histology Structure of SkinDocument4 paginiHistology Structure of SkinLIEBERKHUNÎncă nu există evaluări
- Meth Elab Topic N1Document23 paginiMeth Elab Topic N1Group M1869Încă nu există evaluări
- Skin AnatomyDocument14 paginiSkin Anatomyjellymot3474Încă nu există evaluări
- The Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionDocument14 paginiThe Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionPaolaÎncă nu există evaluări
- General Histology: Unit Three By: Fikre BDocument133 paginiGeneral Histology: Unit Three By: Fikre BHenok GirmaÎncă nu există evaluări
- Lecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalDocument24 paginiLecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalAli Waheed jolan100% (1)
- Skin PDFDocument26 paginiSkin PDFvijendrakumarsahuÎncă nu există evaluări
- EEU2020 Adfinex HDocument23 paginiEEU2020 Adfinex HZainab Jamal SiddiquiÎncă nu există evaluări
- Skin and Soft TissuesDocument122 paginiSkin and Soft TissuesNagulan ChanemougameÎncă nu există evaluări
- Lecture 27 - Histology of The Integumentary System 2Document89 paginiLecture 27 - Histology of The Integumentary System 2spitzmark2030Încă nu există evaluări
- Integumentary SystemDocument24 paginiIntegumentary SystemWadabiÎncă nu există evaluări
- LAB EXERCISE - Fundamental Types of Tissues - Integumentary SystemDocument7 paginiLAB EXERCISE - Fundamental Types of Tissues - Integumentary SystemMickey Ayle De GuzmanÎncă nu există evaluări
- Epithelial Tissue 3-6Document2 paginiEpithelial Tissue 3-6AgungPutraIndraÎncă nu există evaluări
- The Structure and Function of Skin: Section 1Document2 paginiThe Structure and Function of Skin: Section 1MANGÎncă nu există evaluări
- Lesson IntegumentaryDocument31 paginiLesson IntegumentaryTaryhana FitzgerlandÎncă nu există evaluări
- Skin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedDocument10 paginiSkin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedArsene AngelaÎncă nu există evaluări
- 2016 10 18 The Extracellular Matrix (ECM)Document45 pagini2016 10 18 The Extracellular Matrix (ECM)sennaavia12100% (1)
- Tugas Self Directed Learning Sains Dan Biomedik DasarDocument7 paginiTugas Self Directed Learning Sains Dan Biomedik DasarApria PratamaÎncă nu există evaluări
- Skin FasciaDocument16 paginiSkin FasciaManojÎncă nu există evaluări
- Skin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDe la EverandSkin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsÎncă nu există evaluări
- Types of Veneers in Dental WorldDocument6 paginiTypes of Veneers in Dental WorldLenutza LenutaÎncă nu există evaluări
- Randomized Clinical Trial of Indirect Resin Composite and Ceramic Veneers: Up To 3-Year Follow-UpDocument11 paginiRandomized Clinical Trial of Indirect Resin Composite and Ceramic Veneers: Up To 3-Year Follow-UpLenutza LenutaÎncă nu există evaluări
- Critical Review and Evaluation of Compositeceramic Onlaysversus Crowns 2161 1122.1000261Document6 paginiCritical Review and Evaluation of Compositeceramic Onlaysversus Crowns 2161 1122.1000261Lenutza LenutaÎncă nu există evaluări
- Uveitis: Classification, Etiologies and Clinical Signs: E-ISSN 2454-2784Document8 paginiUveitis: Classification, Etiologies and Clinical Signs: E-ISSN 2454-2784Lenutza LenutaÎncă nu există evaluări
- Uveitis: Classification, Etiologies and Clinical Signs: E-ISSN 2454-2784Document8 paginiUveitis: Classification, Etiologies and Clinical Signs: E-ISSN 2454-2784Lenutza LenutaÎncă nu există evaluări
- Studies On The OF Uveitis : Aetiological ProblemDocument12 paginiStudies On The OF Uveitis : Aetiological ProblemLenutza LenutaÎncă nu există evaluări
- Case 62 Chrysler Copr Negotiations Between Diamler and ChryslerDocument46 paginiCase 62 Chrysler Copr Negotiations Between Diamler and ChryslerLenutza LenutaÎncă nu există evaluări
- Wells Score DVTDocument1 paginăWells Score DVTLenutza LenutaÎncă nu există evaluări
- The Histogenesis and Biologic Behavior of Primary Human Malignant Melanomas of The Skin1Document23 paginiThe Histogenesis and Biologic Behavior of Primary Human Malignant Melanomas of The Skin1Lenutza LenutaÎncă nu există evaluări
- Deep Venous ThrombosisDocument22 paginiDeep Venous ThrombosisLenutza LenutaÎncă nu există evaluări
- TopicsDocument1 paginăTopicsLenutza LenutaÎncă nu există evaluări
- NCM 200 - Preparing Administering MedicationDocument21 paginiNCM 200 - Preparing Administering MedicationjhonmarcÎncă nu există evaluări
- Body SymmetryDocument1 paginăBody SymmetryCyanDesÎncă nu există evaluări
- Complications of 3rd Molar SurgeryDocument12 paginiComplications of 3rd Molar Surgeryhamad_kayani_774954Încă nu există evaluări
- MEMORANDU2Document2 paginiMEMORANDU2SanFabian Pangasinan Bfp RegionOneÎncă nu există evaluări
- Artikel Bahasa Inggris (Farmasi)Document3 paginiArtikel Bahasa Inggris (Farmasi)Nadia Putri AuliaÎncă nu există evaluări
- Body PartsDocument4 paginiBody PartsMatthias Dos SantosÎncă nu există evaluări
- MCN II PostpartumDocument16 paginiMCN II PostpartumJharaÎncă nu există evaluări
- Lap Pelayanan SeptemberDocument12 paginiLap Pelayanan SeptemberJosepb SimarmataÎncă nu există evaluări
- ColorgeneticchartangorasDocument7 paginiColorgeneticchartangorasapi-221384858Încă nu există evaluări
- Grammar Workbook Gr. 4 - HomeschoolDocument195 paginiGrammar Workbook Gr. 4 - HomeschoolJessica Pollard Adams100% (2)
- Beating Heart ExplorationDocument1 paginăBeating Heart ExplorationJordan TorresÎncă nu există evaluări
- The.x Files.s01e08.480p.bluray.x264 MRsDocument31 paginiThe.x Files.s01e08.480p.bluray.x264 MRsJohnThorÎncă nu există evaluări
- Hair Analysis Interpretation Guide: The Eck Institute. Issue 27-29Document2 paginiHair Analysis Interpretation Guide: The Eck Institute. Issue 27-29Jianhua Shi100% (1)
- Digestion Absorpton of ProteinsDocument21 paginiDigestion Absorpton of ProteinsSumihar PasaribuÎncă nu există evaluări
- The Nervous System: The Central Processing UnitDocument85 paginiThe Nervous System: The Central Processing UnitGeoffrey MilesÎncă nu există evaluări
- GRADE IV Science QuizDocument2 paginiGRADE IV Science QuizRhea Somollo Bolatin100% (3)
- V. Diagnostic ExaminationDocument3 paginiV. Diagnostic ExaminationArlene MacatangayÎncă nu există evaluări
- Concept Paper Dog SchoolDocument4 paginiConcept Paper Dog SchoolClea Marie Mission100% (2)
- Diagnostic Exam Nepbe All GradesDocument24 paginiDiagnostic Exam Nepbe All GradeselprofeleoÎncă nu există evaluări
- Acid Base AnswersDocument4 paginiAcid Base Answersapi-232466940Încă nu există evaluări
- Fungi & AlgaeDocument3 paginiFungi & AlgaeAulia RizkhanÎncă nu există evaluări
- Flashcards MicrobiologyDocument26 paginiFlashcards MicrobiologyabhishekÎncă nu există evaluări
- LATIHAN SOAL BHS INGGRIS GRADE IX - Peb 25Document12 paginiLATIHAN SOAL BHS INGGRIS GRADE IX - Peb 25Ilham KurniawanÎncă nu există evaluări
- ASDA Released Papers L-MDocument46 paginiASDA Released Papers L-MZuhair Waheed100% (2)
- Path, Rad & NSDocument9 paginiPath, Rad & NSAnonymous MtKJkerbpU0% (1)
- Unit II Cells & Organs of ImmuneDocument69 paginiUnit II Cells & Organs of ImmuneSAMUELÎncă nu există evaluări
- GI Practice Exam With AnswersDocument3 paginiGI Practice Exam With AnswerskcpuffsÎncă nu există evaluări
- Prenatal DevelopmentDocument33 paginiPrenatal DevelopmentDexter Nario0% (1)
- MRCPCH Clinical Exam TechniqueDocument34 paginiMRCPCH Clinical Exam TechniqueRajiv Kabad50% (2)
- Ent Practical GuideDocument176 paginiEnt Practical GuideCiprianRacuciu100% (3)