S-ar putea să vă placă și
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Dolor Postoperatorio y Efectos Secundarios de La Uvulo Palstia Con Radiofrecuencia en Roncopatia Primaria.Document5 paginiDolor Postoperatorio y Efectos Secundarios de La Uvulo Palstia Con Radiofrecuencia en Roncopatia Primaria.Alejandro RuizÎncă nu există evaluări
- The Stony Brook Press - Volume 11, Issue 4Document28 paginiThe Stony Brook Press - Volume 11, Issue 4The Stony Brook PressÎncă nu există evaluări
- Music Recognition, Music Listening, and Word.7Document5 paginiMusic Recognition, Music Listening, and Word.7JIMENEZ PRADO NATALIA ANDREAÎncă nu există evaluări
- Barangay Clearance SampleDocument1 paginăBarangay Clearance SampleBarangay Onse Malaybalay100% (3)
- Narrative Pathophysiology of PregnancyDocument2 paginiNarrative Pathophysiology of PregnancyMarvin ChulyaoÎncă nu există evaluări
- Jis C 8156-2017, EngDocument32 paginiJis C 8156-2017, Eng7620383tlÎncă nu există evaluări
- Air Conditioning Is Inoperative, Fault Code 91eb11 or 91eb15 Is StoredDocument5 paginiAir Conditioning Is Inoperative, Fault Code 91eb11 or 91eb15 Is Storedwasim AiniaÎncă nu există evaluări
- 09B Mechanical Properties of CeramicsDocument13 pagini09B Mechanical Properties of CeramicsAhmed AliÎncă nu există evaluări
- Lecture 8: Separation DesignDocument45 paginiLecture 8: Separation DesignRavi Kiran MÎncă nu există evaluări
- Service Manual - DM0412SDocument11 paginiService Manual - DM0412SStefan Jovanovic100% (1)
- PED16 Foundation of Inclusive Special EducationDocument56 paginiPED16 Foundation of Inclusive Special EducationCHARESS MARSAMOLO TIZONÎncă nu există evaluări
- Hijama Cupping A Review of The EvidenceDocument5 paginiHijama Cupping A Review of The EvidenceDharma Yoga Ayurveda MassageÎncă nu există evaluări
- 15-Statutory Report Statutory Define Law (Legal Protection) Statutory MeetingDocument2 pagini15-Statutory Report Statutory Define Law (Legal Protection) Statutory MeetingRaima DollÎncă nu există evaluări
- Education - Khóa học IELTS 0đ Unit 3 - IELTS FighterDocument19 paginiEducation - Khóa học IELTS 0đ Unit 3 - IELTS FighterAnna TaoÎncă nu există evaluări
- Marantz - dv-4200 DVD Player PDFDocument60 paginiMarantz - dv-4200 DVD Player PDFH.V KayaÎncă nu există evaluări
- Kerala Medico Legal Code - Annexure2Document19 paginiKerala Medico Legal Code - Annexure2doctor82Încă nu există evaluări
- ZX 470Document13 paginiZX 470Mohammed Shaheeruddin100% (1)
- All India Quota Medical Seats (MBBS) With AIPMT 2015 Score PDFDocument5 paginiAll India Quota Medical Seats (MBBS) With AIPMT 2015 Score PDFjuhiÎncă nu există evaluări
- Eim s2000 Series Brochure PDFDocument16 paginiEim s2000 Series Brochure PDFHumbertoOtaloraÎncă nu există evaluări
- Muet Topic 10 City Life Suggested Answer and IdiomsDocument3 paginiMuet Topic 10 City Life Suggested Answer and IdiomsMUHAMAD FAHMI BIN SHAMSUDDIN MoeÎncă nu există evaluări
- R. Raghunandanan - 6Document48 paginiR. Raghunandanan - 6fitrohtin hidayatiÎncă nu există evaluări
- Development of Elevator Ropes: Tech Tip 15Document2 paginiDevelopment of Elevator Ropes: Tech Tip 15أحمد دعبسÎncă nu există evaluări
- CEB 3103 Geotechnical Engineering I: Soil Water and Water FlowDocument39 paginiCEB 3103 Geotechnical Engineering I: Soil Water and Water FlowKia MahiksinhoÎncă nu există evaluări
- Pengaruh Pembangunan Center Point of IndDocument11 paginiPengaruh Pembangunan Center Point of IndSumitro SafiuddinÎncă nu există evaluări
- Quarter 2 Week 5Document54 paginiQuarter 2 Week 5rixzylicoqui.salcedoÎncă nu există evaluări
- Potential Prospect 1.7Document6 paginiPotential Prospect 1.7Joshua M. GenoviaÎncă nu există evaluări
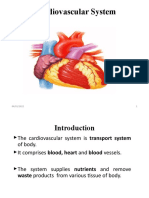
- Cardiovascular SystemDocument47 paginiCardiovascular SystemAtnasiya GadisaÎncă nu există evaluări
- Schengen Certificate ExampleDocument2 paginiSchengen Certificate ExampleGabriel Republi CanoÎncă nu există evaluări
- (PDF) Books Toxic Parents Epub Popular Download - by Susan ForwardDocument1 pagină(PDF) Books Toxic Parents Epub Popular Download - by Susan Forwardmartagonzalezbordonaba0% (3)
- Akin To PityDocument356 paginiAkin To PityPaul StewartÎncă nu există evaluări