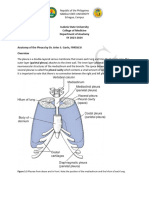
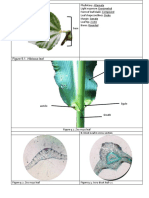
S-ar putea să vă placă și
- Thoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeDe la EverandThoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeÎncă nu există evaluări
- Origin and Development of The Vascular Bundle of Zea Mays L.Document9 paginiOrigin and Development of The Vascular Bundle of Zea Mays L.Afrida NandaÎncă nu există evaluări
- Lab 1Document8 paginiLab 1JeemCarloFagelaPulaÎncă nu există evaluări
- Journal of Entomology and Zoology Volume Eleven, Number Two, June 1919De la EverandJournal of Entomology and Zoology Volume Eleven, Number Two, June 1919Încă nu există evaluări
- New Insights on the Development of the Vascular SystemDe la EverandNew Insights on the Development of the Vascular SystemÎncă nu există evaluări
- Anatomy of Palms R Hap IsDocument22 paginiAnatomy of Palms R Hap IsBelem AlejandroÎncă nu există evaluări
- Stelar System of PlantDocument6 paginiStelar System of Plantworlds tourÎncă nu există evaluări
- Equisetum: The Horsetail PlantDocument7 paginiEquisetum: The Horsetail PlantChithra VijayanÎncă nu există evaluări
- LAB EXERCISE 8 - 10 MM Pig Embryo - Mesodermal DerivativesDocument5 paginiLAB EXERCISE 8 - 10 MM Pig Embryo - Mesodermal DerivativesGerald Angelo DeguinioÎncă nu există evaluări
- Botn 21n - Exercise 12 - Stems and Secondary GrowthDocument17 paginiBotn 21n - Exercise 12 - Stems and Secondary GrowthABIEGAIL LAMBERTÎncă nu există evaluări
- 307 48 ET V1 S1 - File1 PDFDocument22 pagini307 48 ET V1 S1 - File1 PDFaaaatniÎncă nu există evaluări
- Anatomy RenalDocument12 paginiAnatomy RenalAnnisaastikaÎncă nu există evaluări
- Air Tissue Sifton 1945Document36 paginiAir Tissue Sifton 1945Pilar7100Încă nu există evaluări
- Microsurgical Anatomy of Posterior Fossa VeinsDocument43 paginiMicrosurgical Anatomy of Posterior Fossa VeinsCan EkerÎncă nu există evaluări
- The Comparative Morphology of the Winteraceae VI. Vascular Anatomy of the Flowering ShootDocument18 paginiThe Comparative Morphology of the Winteraceae VI. Vascular Anatomy of the Flowering Shootbarerahjaved479Încă nu există evaluări
- The Origin and Conduction of The Heart BeatDocument43 paginiThe Origin and Conduction of The Heart BeatDjanino Fernandes0% (1)
- Un Estudio de TargioniaDocument13 paginiUn Estudio de TargioniaramongonzaÎncă nu există evaluări
- Strucrtures of Plant PartsDocument95 paginiStrucrtures of Plant Partsfatima100% (1)
- On A New Organ in The Lycoridea, and On The Nephridium in Nereis Diversicolor, O. P. MullDocument18 paginiOn A New Organ in The Lycoridea, and On The Nephridium in Nereis Diversicolor, O. P. Mullelfan titiansyahÎncă nu există evaluări
- Young Stems and RootsDocument11 paginiYoung Stems and RootsHailey CoutuÎncă nu există evaluări
- Plant Physiology LAB REPORT XDocument5 paginiPlant Physiology LAB REPORT XBoyd benson kayomboÎncă nu există evaluări
- Anatomy and Properties of Bamboo CulmsDocument198 paginiAnatomy and Properties of Bamboo Culmsvin_3ldÎncă nu există evaluări
- Vascular Tissue DifferentiationDocument9 paginiVascular Tissue DifferentiationC'estMoiÎncă nu există evaluări
- Fig. 2 B, (Fig. at Of: Histology EyeDocument26 paginiFig. 2 B, (Fig. at Of: Histology EyeraniaÎncă nu există evaluări
- Pteridophytes-Telome Theory and Stelar Evolution by Dr. Prabha DhondiyalDocument39 paginiPteridophytes-Telome Theory and Stelar Evolution by Dr. Prabha DhondiyalAbhishek Singh ChandelÎncă nu există evaluări
- The Vascular Endothelium: A Holistic Approach for OncologyDe la EverandThe Vascular Endothelium: A Holistic Approach for OncologyÎncă nu există evaluări
- NHLBI Workshop Summary: The Mysterious Pulmonary Brush CellDocument4 paginiNHLBI Workshop Summary: The Mysterious Pulmonary Brush CellcholoconvertiÎncă nu există evaluări
- ST Joseph's Institution (Senior School) IBDP1 HL Biology PracticalDocument5 paginiST Joseph's Institution (Senior School) IBDP1 HL Biology PracticalSean NgÎncă nu există evaluări
- Top 3 Shoot Apical Meristem TheoriesDocument11 paginiTop 3 Shoot Apical Meristem TheoriesAbhimanyu Pandey100% (2)
- Theories of Shoot Apical Meristem OKOKOKDocument8 paginiTheories of Shoot Apical Meristem OKOKOKShameel PervezÎncă nu există evaluări
- A4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Document16 paginiA4. Thilander H, Bloom G. Cell Contacts in Oral Epithelia. J Perio Res 1968Daniel CubasÎncă nu există evaluări
- The Biology of Lumbriculus variegatusDocument4 paginiThe Biology of Lumbriculus variegatusklipschthx0Încă nu există evaluări
- Carl Claus - XIII.-On The Organization of The CypridesDocument5 paginiCarl Claus - XIII.-On The Organization of The CypridestorigÎncă nu există evaluări
- Chapter 140: Cochlear Anatomy and Central Auditory Pathways Peter A. Santi, Patrizia ManciniDocument20 paginiChapter 140: Cochlear Anatomy and Central Auditory Pathways Peter A. Santi, Patrizia ManciniJustkillme AlifÎncă nu există evaluări
- Senecio Grandidentatus Ledeb. (Asteraceae) : Anatomical Aspects of The Stem and Leaf ofDocument7 paginiSenecio Grandidentatus Ledeb. (Asteraceae) : Anatomical Aspects of The Stem and Leaf ofBlinda Raluca MariaÎncă nu există evaluări
- Mephitinve, Spilogale, Conepatus, Me Phitis. Mephgtis Conetatus. Mefhlt S: M. Macrura, .M. 2utorlus, MethitlcaDocument18 paginiMephitinve, Spilogale, Conepatus, Me Phitis. Mephgtis Conetatus. Mefhlt S: M. Macrura, .M. 2utorlus, MethitlcaLouis CyferÎncă nu există evaluări
- Anatomia Foramem IntervertebralDocument11 paginiAnatomia Foramem IntervertebralmlmudoÎncă nu există evaluări
- Gross Anatomy II Lecture on Heart and Vascular SystemDocument13 paginiGross Anatomy II Lecture on Heart and Vascular SystemSantosh Bhandari100% (2)
- Journal of Morphology SpermwarteryDocument14 paginiJournal of Morphology SpermwarteryJustin RamonÎncă nu există evaluări
- BOTANYDocument5 paginiBOTANYPrincess Fay LopezÎncă nu există evaluări
- PleuraDocument6 paginiPleurasjs6r8wwv9Încă nu există evaluări
- Jurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas ADocument17 paginiJurnal - Christopher Muhamad Zildan Holliday - 215090107111029 - Kelas AChristopher Muhamad Zildan HÎncă nu există evaluări
- The Stem and Root Anatomy of Sanmiguelia lewisiiDocument41 paginiThe Stem and Root Anatomy of Sanmiguelia lewisiiShafa NnisaÎncă nu există evaluări
- Miología Apendicular Del Procyon Cancrivorus (Windle, 1888)Document9 paginiMiología Apendicular Del Procyon Cancrivorus (Windle, 1888)Sebastian ChungaÎncă nu există evaluări
- Lecture Learning Objectives for BIO 210 SystemsDocument14 paginiLecture Learning Objectives for BIO 210 SystemsAmber DavisÎncă nu există evaluări
- Ventrilces BrainDocument18 paginiVentrilces BrainVamsi KrishnaÎncă nu există evaluări
- Human Anat 2 Year Volume 4Document122 paginiHuman Anat 2 Year Volume 4HARSHINI PANDIANÎncă nu există evaluări
- VeinsDocument5 paginiVeinsMaximos ManiatisÎncă nu există evaluări
- NyctaDocument18 paginiNyctaAghila SamjiÎncă nu există evaluări
- Tissue and Tissue Systems - STD 9Document14 paginiTissue and Tissue Systems - STD 9Syamala Natarajan100% (1)
- Blood Vessels, AnatomyDocument20 paginiBlood Vessels, AnatomyAbu BakarÎncă nu există evaluări
- ANATOMICAL STUDY OF ANOMALY STRUCTURE IN CucurbitaDocument5 paginiANATOMICAL STUDY OF ANOMALY STRUCTURE IN CucurbitaGaluh Novita Ayu0% (1)
- Chapter 6 Anatomy of Flowering PlantsDocument8 paginiChapter 6 Anatomy of Flowering Plantsswayamdey71Încă nu există evaluări
- Life Cycle and Reproduction of Anthoceros LiverwortDocument24 paginiLife Cycle and Reproduction of Anthoceros LiverwortListiyani Putri AzzahroÎncă nu există evaluări
- Equisetum 180922130237Document34 paginiEquisetum 180922130237Ján ŠormanÎncă nu există evaluări
- Structure: Plant Form and FunctionDocument12 paginiStructure: Plant Form and FunctionPhilippines QatarÎncă nu există evaluări
- Botany I PDFDocument4 paginiBotany I PDFFakiha NadeemÎncă nu există evaluări
- Tamil Nadu 11th Standard Botany Practical ManualDocument43 paginiTamil Nadu 11th Standard Botany Practical Manualsri thamizhan80% (85)
- Northern beech trunk in autumnDocument4 paginiNorthern beech trunk in autumnmansi bavliyaÎncă nu există evaluări
- Exercise 7. Leaf: Distinctive CharacteristicsDocument8 paginiExercise 7. Leaf: Distinctive CharacteristicsTers MedinaÎncă nu există evaluări
- PLANTS: Types, Parts, Uses & How They Make FoodDocument22 paginiPLANTS: Types, Parts, Uses & How They Make FoodSurbhi Bhatia100% (1)
- Aquatic Plants AdaptDocument2 paginiAquatic Plants AdaptKrystallane ManansalaÎncă nu există evaluări
- Transport System in PlantDocument16 paginiTransport System in PlantAbu-Omar50% (4)
- Practical: I Total Marks 50 Five Questions Will Be Asked Each Question Carries 10 MarksDocument3 paginiPractical: I Total Marks 50 Five Questions Will Be Asked Each Question Carries 10 MarksCrazy GamerÎncă nu există evaluări
- Identify The Plant Organ ActivityDocument5 paginiIdentify The Plant Organ Activityapi-246973610Încă nu există evaluări
- 1 - Morphology of The Rice PlantDocument31 pagini1 - Morphology of The Rice PlantThe Municipal Agriculturist100% (1)
- Morphology and Anatomy of Flowering PlantsDocument3 paginiMorphology and Anatomy of Flowering PlantsAyako HayashidaÎncă nu există evaluări
- Vegetative OrgansDocument68 paginiVegetative OrgansAngel GoÎncă nu există evaluări
- Exer 9 LeavesDocument3 paginiExer 9 LeavesColleen Dejan100% (1)
- Struktur Anatomi Batang Tiga Jenis Rotan Genus Calamus Dari Kawasan Tembawang Kabupaten Kubu RayaDocument6 paginiStruktur Anatomi Batang Tiga Jenis Rotan Genus Calamus Dari Kawasan Tembawang Kabupaten Kubu RayaZahra Nur AzizahÎncă nu există evaluări
- AnfistumDocument207 paginiAnfistumjosefannyhahaÎncă nu există evaluări
- ELSC Week13 Plant Survival PDFDocument43 paginiELSC Week13 Plant Survival PDFarriane BaladadÎncă nu există evaluări
- Exercise 2Document7 paginiExercise 2Lendie VillÎncă nu există evaluări
- CBSE Class 11 Biology Chapter 6 Anatomy of Flowering Plants Revision NotesDocument72 paginiCBSE Class 11 Biology Chapter 6 Anatomy of Flowering Plants Revision Noteschakrabortytrisha.2006Încă nu există evaluări
- Bryanston Primary School: Natural Science Cycle TestDocument8 paginiBryanston Primary School: Natural Science Cycle TestErinÎncă nu există evaluări
- Chapter 1 (Part 1) - Plant Anatomy & Physiology PDFDocument89 paginiChapter 1 (Part 1) - Plant Anatomy & Physiology PDFMohammed Ridzuwan94% (16)
- NEET Level Test Questions (29-Jan-23Document25 paginiNEET Level Test Questions (29-Jan-23Unit VIIÎncă nu există evaluări
- Modul Perfect Score SBP Biology SPM 2013 Question and SchemeDocument283 paginiModul Perfect Score SBP Biology SPM 2013 Question and SchemeCikgu Faizal87% (15)
- Cells-Tissues Student GuideDocument2 paginiCells-Tissues Student GuideMark LozoÎncă nu există evaluări
- Plant Body Plant Body: Why Is Histology Important?Document6 paginiPlant Body Plant Body: Why Is Histology Important?ZyreeneNicoleÎncă nu există evaluări
- Plant Cell Specialization and Tissue DifferentiationDocument10 paginiPlant Cell Specialization and Tissue DifferentiationAndrea CruzÎncă nu există evaluări
- 8384 ST PDFDocument20 pagini8384 ST PDFJohn Dave Francisco100% (1)
- Bio MaterialsDocument26 paginiBio MaterialsGeorge SingerÎncă nu există evaluări
- Anatomy of Flowering PlantsDocument19 paginiAnatomy of Flowering PlantsPrerak PatelÎncă nu există evaluări
- CucurbitaDocument5 paginiCucurbitaapi-49891619Încă nu există evaluări