S-ar putea să vă placă și
- BIO-Resonance Diagnostic Systems OBERON - Flyer v2Document3 paginiBIO-Resonance Diagnostic Systems OBERON - Flyer v2OBERON-INTROSPECT-BIOSPECTÎncă nu există evaluări
- Tail PCR ProtocolDocument4 paginiTail PCR ProtocolFelipe ZapataÎncă nu există evaluări
- LymphomaDocument78 paginiLymphomaastriwahyuni100% (4)
- Caillaud Et Al 2008 Sex Selection Morpho RencDocument10 paginiCaillaud Et Al 2008 Sex Selection Morpho Rencferet33947Încă nu există evaluări
- Identifying Individual Ungulates From Fecal Dna A Comparison of Field Collection Methods To Maximize Efficiency, Ease, and SuccessDocument12 paginiIdentifying Individual Ungulates From Fecal Dna A Comparison of Field Collection Methods To Maximize Efficiency, Ease, and SuccessFah NichapatÎncă nu există evaluări
- Predicting The Sex of Dovekies by Discriminant AnalysisDocument6 paginiPredicting The Sex of Dovekies by Discriminant AnalysisIvan JerkovićÎncă nu există evaluări
- Molecular Genetic Analysis of Male Alternative Strategy and Reproductive Success in The Polygynous Mating BatDocument9 paginiMolecular Genetic Analysis of Male Alternative Strategy and Reproductive Success in The Polygynous Mating BatSiti Rabiatul FajriÎncă nu există evaluări
- Ancient Dna To Decipher The Domestication of DogDocument6 paginiAncient Dna To Decipher The Domestication of DogIvan KlarićÎncă nu există evaluări
- 1 s2.0 S266693742100024X MainDocument7 pagini1 s2.0 S266693742100024X Mainitalabezerra54Încă nu există evaluări
- JZO 2020 Tourani EtalDocument13 paginiJZO 2020 Tourani EtalJoana FerreiraÎncă nu există evaluări
- Meza Oviedo2020Document5 paginiMeza Oviedo2020Ljón BjörnÎncă nu există evaluări
- Biology, Ecology and Systematics of Australian Scelio: Wasp Parasitoids of Locust and Grasshopper EggsDe la EverandBiology, Ecology and Systematics of Australian Scelio: Wasp Parasitoids of Locust and Grasshopper EggsÎncă nu există evaluări
- Grooming EnglishDocument13 paginiGrooming Englishluisa fernanda jimenez gutierrezÎncă nu există evaluări
- UrineDocument11 paginiUrineShubham PatwaÎncă nu există evaluări
- Experimental Study of Bone Modification by Captive CaracalDocument13 paginiExperimental Study of Bone Modification by Captive CaracalSidra ZahoorÎncă nu există evaluări
- Detection Distance and Environmental Factors in CoDocument11 paginiDetection Distance and Environmental Factors in Cokenanpa7590Încă nu există evaluări
- Kery 2002Document10 paginiKery 2002Reyna GutiérrezÎncă nu există evaluări
- Ticks and Tick-Borne DiseasesDocument8 paginiTicks and Tick-Borne DiseasesLørïïs SïerraÎncă nu există evaluări
- Ecosphere - 2021 - Doran Myers - Density Estimates For Canada Lynx Vary Among Estimation MethodsDocument18 paginiEcosphere - 2021 - Doran Myers - Density Estimates For Canada Lynx Vary Among Estimation MethodsDennis MurrayÎncă nu există evaluări
- Baker & Bradley 2006. Speciation in Mammals and The Genetic Species ConceptDocument20 paginiBaker & Bradley 2006. Speciation in Mammals and The Genetic Species ConceptpagomezsÎncă nu există evaluări
- Detection Dogs Allow For Systematic Non-Invasive Collection of DNA Samples From Eurasian LynxDocument5 paginiDetection Dogs Allow For Systematic Non-Invasive Collection of DNA Samples From Eurasian Lynxsusey madelit apaza mamaniÎncă nu există evaluări
- Wa0000.Document76 paginiWa0000.kolkarevinayÎncă nu există evaluări
- Cozzi2021 Article EyesEarsOrNoseComparisonOfThreDocument13 paginiCozzi2021 Article EyesEarsOrNoseComparisonOfThreRossana AstolfiÎncă nu există evaluări
- Ap Genetic Diversity Inbreeding and Breeding Practices in Dogs Resultsfrom Pedigree AnalysesDocument6 paginiAp Genetic Diversity Inbreeding and Breeding Practices in Dogs Resultsfrom Pedigree Analysesapi-458387634Încă nu există evaluări
- tmp34B9 TMPDocument9 paginitmp34B9 TMPFrontiersÎncă nu există evaluări
- Veterinary Record - 2016 - Pugh - Cumulative Incidence and Risk Factors For Limber Tail in The Dogslife Labrador RetrieverDocument5 paginiVeterinary Record - 2016 - Pugh - Cumulative Incidence and Risk Factors For Limber Tail in The Dogslife Labrador RetrieverZullyÎncă nu există evaluări
- Fevo 10 910695Document13 paginiFevo 10 910695Ayane BastosÎncă nu există evaluări
- Reppucci Et Al 2011Document10 paginiReppucci Et Al 2011Juan ReppucciÎncă nu există evaluări
- 红狼和土狼杂交Document12 pagini红狼和土狼杂交王刚Încă nu există evaluări
- 2020 Saproxylic Cetoniidae Coleoptera SCDocument8 pagini2020 Saproxylic Cetoniidae Coleoptera SCJuan José Sánchez MecaÎncă nu există evaluări
- Forensic EvidenceDocument36 paginiForensic EvidenceBryan MagcantaÎncă nu există evaluări
- Tracking Cryptic Animals KershenbaumDocument12 paginiTracking Cryptic Animals KershenbaumJorge Caro BernalÎncă nu există evaluări
- Sex and Weaponry: The Distribution of Toxin-Storage Glands On The Bodies of Male and Female Cane Toads (Rhinella Marina)Document8 paginiSex and Weaponry: The Distribution of Toxin-Storage Glands On The Bodies of Male and Female Cane Toads (Rhinella Marina)cooki rumiyahÎncă nu există evaluări
- Novel North American Hominins Final PDF DownloadDocument41 paginiNovel North American Hominins Final PDF DownloadRichard Bachman100% (1)
- CONECCIÓN PERRO HUMANO Ar.24534Document9 paginiCONECCIÓN PERRO HUMANO Ar.24534Lisette Velásquez AnticonaÎncă nu există evaluări
- Nagataetal 2022MammalStudyDocument10 paginiNagataetal 2022MammalStudyNealÎncă nu există evaluări
- Preproposal FinalDocument9 paginiPreproposal Finalapi-254424729Încă nu există evaluări
- Freeb Honors ReseachpDocument13 paginiFreeb Honors Reseachpapi-701058724Încă nu există evaluări
- Proposal To Sequence The First Reptilian Genome: The Green Anole Lizard, Anolis CarolinensisDocument21 paginiProposal To Sequence The First Reptilian Genome: The Green Anole Lizard, Anolis CarolinensisJamesComeyJustaBitchÎncă nu există evaluări
- Statement of The ProblemDocument10 paginiStatement of The ProblemCyril D. SuazoÎncă nu există evaluări
- New Glider SpeciesDocument5 paginiNew Glider Specieskim haroldÎncă nu există evaluări
- Krueger Etal 12Document30 paginiKrueger Etal 12Juan David LozanoÎncă nu există evaluări
- Alibhai The Challenge of Monitoring Elusive LaDocument22 paginiAlibhai The Challenge of Monitoring Elusive LaCippalippaÎncă nu există evaluări
- Developments in Amphibian Parental Care Research: History, Present Advances, and Future PerspectivesDocument27 paginiDevelopments in Amphibian Parental Care Research: History, Present Advances, and Future PerspectivesMarlonAndrésPovedaSánchezÎncă nu există evaluări
- Id KezDocument129 paginiId KezAxi AxiÎncă nu există evaluări
- Albinism SpydermonkeysDocument8 paginiAlbinism SpydermonkeysAnibal Silva PioliÎncă nu există evaluări
- Ward Et Al 2009 - The Campaign To DNA Barcode All FishesDocument28 paginiWard Et Al 2009 - The Campaign To DNA Barcode All FishesEduardo TLÎncă nu există evaluări
- Morphology of Male and Female ReproductiveDocument12 paginiMorphology of Male and Female ReproductivemariarosiemillybioÎncă nu există evaluări
- Scientists Solve The Riddle of Zebras' StripesDocument2 paginiScientists Solve The Riddle of Zebras' StripesSyma ShaheenÎncă nu există evaluări
- Assisting Decision-Making On Age of Neutering For 35 Breeds of Dogs: Associated Joint Disorders, Cancers, and Urinary IncontinenceDocument14 paginiAssisting Decision-Making On Age of Neutering For 35 Breeds of Dogs: Associated Joint Disorders, Cancers, and Urinary IncontinenceTaires RodriguesÎncă nu există evaluări
- Research Paper On SquirrelsDocument9 paginiResearch Paper On Squirrelslqinlccnd100% (1)
- Achievements in The Life Sciences: Vitaliy A. Zaitsev, Ivan V. Seryodkin, Dariya A. Maksimova, Svetlana V. SoutyrinaDocument4 paginiAchievements in The Life Sciences: Vitaliy A. Zaitsev, Ivan V. Seryodkin, Dariya A. Maksimova, Svetlana V. Soutyrinasameer sahaanÎncă nu există evaluări
- Zhang 2011Document9 paginiZhang 2011Enrique AlvaradoÎncă nu există evaluări
- 2016 - A Comparison of Animal Jaws and Bite Mark PatternsDocument15 pagini2016 - A Comparison of Animal Jaws and Bite Mark Patternsconstanza.salazar.silvaÎncă nu există evaluări
- Eat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsDocument5 paginiEat Your Heart Out: Choice and Handling of Novel Toxic Prey by Predatory Water RatsRudson RomeroÎncă nu există evaluări
- Meiklejohn Et Al 2021Document25 paginiMeiklejohn Et Al 2021JudicaëlÎncă nu există evaluări
- Kaiser Et Al. 2016 Mer PDFDocument11 paginiKaiser Et Al. 2016 Mer PDFAditya WirawanÎncă nu există evaluări
- Patterns of Praying Mantis Auditory System EvolutiDocument29 paginiPatterns of Praying Mantis Auditory System EvolutiromyÎncă nu există evaluări
- Eat Your Heart Out - Choice and Handling of Novel Toxic Prey by Predatory Water RatsDocument6 paginiEat Your Heart Out - Choice and Handling of Novel Toxic Prey by Predatory Water RatsJean Carlos MirandaÎncă nu există evaluări
- OrqaDocument10 paginiOrqaTulio EnríquezÎncă nu există evaluări
- Salaetal 2014QIwildandcaptivewolfmodificationsDocument11 paginiSalaetal 2014QIwildandcaptivewolfmodificationsangelthaiboxyahoo.esÎncă nu există evaluări
- A Generalised Index ASSIS 1996Document5 paginiA Generalised Index ASSIS 1996Lucía SolerÎncă nu există evaluări
- A New Era For Carnivore ConservationDocument5 paginiA New Era For Carnivore ConservationLucía SolerÎncă nu există evaluări
- 464 FullDocument11 pagini464 FullLucía SolerÎncă nu există evaluări
- Prioritizing Conservation Investments For Mammal Species GloballyDocument11 paginiPrioritizing Conservation Investments For Mammal Species GloballyLucía SolerÎncă nu există evaluări
- Bernal Bravo, César Angulo Rasco, Félix Interacciones de Los Jóvenes Andaluces en Las Redes Sociales Comunicar, Vol. XX, Núm. 40, Marzo-Octubre, 2013, Pp. 25-30 Grupo Comunicar Huelva, EspañaDocument7 paginiBernal Bravo, César Angulo Rasco, Félix Interacciones de Los Jóvenes Andaluces en Las Redes Sociales Comunicar, Vol. XX, Núm. 40, Marzo-Octubre, 2013, Pp. 25-30 Grupo Comunicar Huelva, EspañaLucía SolerÎncă nu există evaluări
- Challenges of Large Carnivore Conservation: Sloth Bear Attacks in Sri LankaDocument14 paginiChallenges of Large Carnivore Conservation: Sloth Bear Attacks in Sri LankaLucía SolerÎncă nu există evaluări
- Vucetich J A Et Al 2018 Just Conservation What Is It and Should We Pursue ItDocument11 paginiVucetich J A Et Al 2018 Just Conservation What Is It and Should We Pursue ItLucía SolerÎncă nu există evaluări
- Art Cat CastDocument17 paginiArt Cat CastLucía SolerÎncă nu există evaluări
- 5 Critical Exploration in The University Classroom PDFDocument9 pagini5 Critical Exploration in The University Classroom PDFLucía SolerÎncă nu există evaluări
- Carvalho Et Al 2012 PDFDocument14 paginiCarvalho Et Al 2012 PDFLucía SolerÎncă nu există evaluări
- Art Cat CastDocument17 paginiArt Cat CastLucía SolerÎncă nu există evaluări
- Arboretum Images 1Document8 paginiArboretum Images 1Lucía SolerÎncă nu există evaluări
- A Sense of Mystery - The Scientist Magazine®Document12 paginiA Sense of Mystery - The Scientist Magazine®Lucía Soler100% (1)
- Tesis - Ditmar KurtzDocument110 paginiTesis - Ditmar KurtzLucía SolerÎncă nu există evaluări
- The Role of Human Dimensions in Wildlife Resource Research IN Wildlife ManagementDocument7 paginiThe Role of Human Dimensions in Wildlife Resource Research IN Wildlife ManagementLucía SolerÎncă nu există evaluări
- Palm Community Transitions Along A Topographic Gradient From Floodplain To Terra Firme in The Eastern AmazonDocument10 paginiPalm Community Transitions Along A Topographic Gradient From Floodplain To Terra Firme in The Eastern AmazonLucía SolerÎncă nu există evaluări
- Di Marco M 2014 Drivers of Extinction Risk in African Mammals The Interplay of Distribution State Human Pressure Conservation Response and Species BiologyDocument12 paginiDi Marco M 2014 Drivers of Extinction Risk in African Mammals The Interplay of Distribution State Human Pressure Conservation Response and Species BiologyLucía SolerÎncă nu există evaluări
- Geo-Spatial Aspects of Acceptance of Illegal Hunting of Large Carnivores in ScandinaviaDocument9 paginiGeo-Spatial Aspects of Acceptance of Illegal Hunting of Large Carnivores in ScandinaviaLucía SolerÎncă nu există evaluări
- A Knowledge-Based Approach To Estimating The Magnitude and Spatial 2016Document10 paginiA Knowledge-Based Approach To Estimating The Magnitude and Spatial 2016Lucía SolerÎncă nu există evaluări
- 2015 - Robbins Et Al - Political Ecology - Conservation LettersDocument9 pagini2015 - Robbins Et Al - Political Ecology - Conservation LettersLucía SolerÎncă nu există evaluări
- Cianfrani AniCon2013Document11 paginiCianfrani AniCon2013Lucía SolerÎncă nu există evaluări
- 35 79 1 SMDocument26 pagini35 79 1 SMLucía SolerÎncă nu există evaluări
- CN58 Kalle Et Al - Cat NewsDocument12 paginiCN58 Kalle Et Al - Cat NewsLucía SolerÎncă nu există evaluări
- Conservation of Carnivore Species in West AfricaDocument30 paginiConservation of Carnivore Species in West AfricaLucía SolerÎncă nu există evaluări
- 12 - Chapter 4 Prey Selection and Food Habits of LargeDocument38 pagini12 - Chapter 4 Prey Selection and Food Habits of LargeLucía SolerÎncă nu există evaluări
- Disponibilidade de Frutos e A Dieta de Lycalopex Vetulus (Carnivora - Canidae) em Um Cerrado de Mato Grosso, BrasilDocument8 paginiDisponibilidade de Frutos e A Dieta de Lycalopex Vetulus (Carnivora - Canidae) em Um Cerrado de Mato Grosso, BrasilLucía SolerÎncă nu există evaluări
- Naves Cons Biol 03Document14 paginiNaves Cons Biol 03Lucía SolerÎncă nu există evaluări
- ThreatsFishBiodiversity PDFDocument12 paginiThreatsFishBiodiversity PDFLucía SolerÎncă nu există evaluări
- 09 08 10 - 266 281 - Lapenta ProcopioDocument16 pagini09 08 10 - 266 281 - Lapenta ProcopioLucía SolerÎncă nu există evaluări
- Introductory Paper On Critical Explorations in Teaching Art, Science, and Teacher EducationDocument16 paginiIntroductory Paper On Critical Explorations in Teaching Art, Science, and Teacher EducationLucía SolerÎncă nu există evaluări
- Chapter 1.3 Structures and Functions of OrganellesDocument35 paginiChapter 1.3 Structures and Functions of OrganellesFairview PenangÎncă nu există evaluări
- Year 10 DPP Biology Term 3 Exam RevisionDocument8 paginiYear 10 DPP Biology Term 3 Exam RevisionPoojit VenugopalaÎncă nu există evaluări
- Antibacterial activities of the methanol extracts of Albizia adianthifolia, Alchornea laxiflora, Laportea ovalifolia and three other Cameroonian plants against multi-drug resistant Gram-negative bacteriaDocument6 paginiAntibacterial activities of the methanol extracts of Albizia adianthifolia, Alchornea laxiflora, Laportea ovalifolia and three other Cameroonian plants against multi-drug resistant Gram-negative bacteriaArieÎncă nu există evaluări
- Anatomi TumbuhanDocument30 paginiAnatomi TumbuhanMEISYI DSÎncă nu există evaluări
- Cytogenetics Transes Topic 16Document5 paginiCytogenetics Transes Topic 16Dianne NolascoÎncă nu există evaluări
- Hiv Associated Hematological Malignasis - Marcus HentrichDocument239 paginiHiv Associated Hematological Malignasis - Marcus HentrichJaronjer1Încă nu există evaluări
- Telescopium DewasaDocument11 paginiTelescopium DewasaBayu KrisnaÎncă nu există evaluări
- Alzheimers-Disease-Genetics-Fact-Sheet 0Document8 paginiAlzheimers-Disease-Genetics-Fact-Sheet 0api-285676076Încă nu există evaluări
- Effective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsDocument19 paginiEffective Microbial Bioremediation Via The Multi-Omics Approach: An Overview of Trends, Problems and ProspectsUMYU Journal of Microbiology Research (UJMR)Încă nu există evaluări
- Lecture D111L Week 05 S13Document64 paginiLecture D111L Week 05 S13Tahir RizviÎncă nu există evaluări
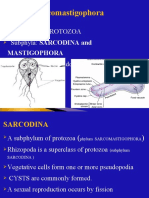
- Sarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or PseudopodiaDocument20 paginiSarcomastigophora: A Phylum of PROTOZOA Subphyla: SARCODINA and Have Flagella or Pseudopodiaabel semuÎncă nu există evaluări
- 2011 Fractal DNADocument7 pagini2011 Fractal DNAAbby SweeneyÎncă nu există evaluări
- Lecture 1Document16 paginiLecture 1PappuÎncă nu există evaluări
- NematodeschapterDocument9 paginiNematodeschapterArjelyn Mae D BuendiaÎncă nu există evaluări
- Prescotts Microbiology 10th Edition Willey Solutions Manual 1Document4 paginiPrescotts Microbiology 10th Edition Willey Solutions Manual 1ruth98% (48)
- Bacteriophage Replication ProcessDocument3 paginiBacteriophage Replication ProcessNicole JulianÎncă nu există evaluări
- 1-17 Microbiology Lab ApparatusDocument2 pagini1-17 Microbiology Lab ApparatusNathaniel PulidoÎncă nu există evaluări
- Nature at Work - The Ongoing Saga of Evolution - 400 PagsDocument400 paginiNature at Work - The Ongoing Saga of Evolution - 400 PagsNoti News WorldÎncă nu există evaluări
- Cmi CMBDocument7 paginiCmi CMBMary M. ToribioÎncă nu există evaluări
- BIOLOGY Form 5 Chapter 1 TransportDocument42 paginiBIOLOGY Form 5 Chapter 1 TransportShephard Png91% (95)
- (Biology) The LiverDocument7 pagini(Biology) The LiverNiharikaÎncă nu există evaluări
- Tuesday, August 18, 2009Document47 paginiTuesday, August 18, 2009zorbaxÎncă nu există evaluări
- Pneumacult™-Ali: For Bronchial Epithelial CellsDocument2 paginiPneumacult™-Ali: For Bronchial Epithelial CellsDunkMeÎncă nu există evaluări
- Plant Symbionts InteractionDocument13 paginiPlant Symbionts InteractionIram TahirÎncă nu există evaluări
- Obesity 1. Basic Science of ObesityDocument10 paginiObesity 1. Basic Science of ObesitygraceÎncă nu există evaluări
- Anatomy& Physiology BY Adeniyi M. OlatoyeDocument59 paginiAnatomy& Physiology BY Adeniyi M. OlatoyeAdeniyi OlatoyeÎncă nu există evaluări
- Lecture#1: Plant Cell and Tissue CultureDocument12 paginiLecture#1: Plant Cell and Tissue CultureIbrahimAslamÎncă nu există evaluări