S-ar putea să vă placă și
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Housestaff Manual 2023-2024 - MeadeDocument267 paginiHousestaff Manual 2023-2024 - MeadeMelchor Alcántara Barrera100% (2)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Principle of Animal Physiology (Christopher D. Moyes, Patricia Schulte) Principle ofDocument763 paginiPrinciple of Animal Physiology (Christopher D. Moyes, Patricia Schulte) Principle ofKing Amv86% (7)
- Chest PainDocument31 paginiChest PainMarwan GamaleldinÎncă nu există evaluări
- Human Anatomy and Physiology MCQ With Answers PDFDocument4 paginiHuman Anatomy and Physiology MCQ With Answers PDFPrince Maurya88% (8)
- Patent Ductus Arteriosus (PDA) Diagnosis and TreatmentDocument19 paginiPatent Ductus Arteriosus (PDA) Diagnosis and Treatmentputri avriantiÎncă nu există evaluări
- Republic Act 3019 (RA 3019) - Anti-Graft LawDocument5 paginiRepublic Act 3019 (RA 3019) - Anti-Graft LawJun RinonÎncă nu există evaluări
- Fitness, Sports and Recreation Leadership: Quarter 3 - Module 2Document26 paginiFitness, Sports and Recreation Leadership: Quarter 3 - Module 2Regie Carino100% (2)
- L3 A&p#3 With AnswersDocument7 paginiL3 A&p#3 With AnswersFergus DaleÎncă nu există evaluări
- IIT/FIELD MUSEUM High School Lab on Plant PigmentsDocument4 paginiIIT/FIELD MUSEUM High School Lab on Plant PigmentsGeraldineMay50% (2)
- Ecol483 Lab 12 - Reptile Anatomy PDFDocument16 paginiEcol483 Lab 12 - Reptile Anatomy PDFGeraldineMayÎncă nu există evaluări
- BIOCHEM LAB - Enzymatic Activity of Salivary AmylaseDocument6 paginiBIOCHEM LAB - Enzymatic Activity of Salivary AmylaseIsmael Cortez100% (2)
- Rizal in Ateneo de Manila and USTDocument8 paginiRizal in Ateneo de Manila and USTGeraldineMayÎncă nu există evaluări
- CirclesDocument3 paginiCirclesGeraldineMayÎncă nu există evaluări
- Right To Conscience JournalDocument8 paginiRight To Conscience JournalGeraldineMayÎncă nu există evaluări
- IplanelabDocument9 paginiIplanelabromwamaÎncă nu există evaluări
- Gilbert Villanueva Grade 9 StudentDocument1 paginăGilbert Villanueva Grade 9 StudentGeraldineMayÎncă nu există evaluări
- Questions To AnswerDocument1 paginăQuestions To AnswerGeraldineMayÎncă nu există evaluări
- Physics Friction LabDocument7 paginiPhysics Friction LabGeraldineMayÎncă nu există evaluări
- Spinal ReflexDocument6 paginiSpinal ReflexGeraldineMayÎncă nu există evaluări
- Greek CivilizationDocument2 paginiGreek CivilizationGeraldineMayÎncă nu există evaluări
- Abiotic and LocationDocument1 paginăAbiotic and LocationGeraldineMayÎncă nu există evaluări
- Survey ResultsDocument15 paginiSurvey ResultsGeraldineMayÎncă nu există evaluări
- Kinetic Friction Experiment Finds Surface Material Affects CoefficientDocument8 paginiKinetic Friction Experiment Finds Surface Material Affects Coefficientkate anne del castroÎncă nu există evaluări
- Improving Mental HealthDocument11 paginiImproving Mental HealthGeraldineMayÎncă nu există evaluări
- FrictionDocument2 paginiFrictionGeraldineMayÎncă nu există evaluări
- Rizal LawDocument5 paginiRizal LawMathilda Victoria GomezÎncă nu există evaluări
- Promissory Note SampleDocument1 paginăPromissory Note SampleGeraldineMayÎncă nu există evaluări
- ENZYMESDocument57 paginiENZYMESGeraldineMayÎncă nu există evaluări
- Enzymes in Action: Understanding Factors that Affect Enzyme ActivityDocument15 paginiEnzymes in Action: Understanding Factors that Affect Enzyme Activitysung_kei_pinÎncă nu există evaluări
- Amylase LabDocument2 paginiAmylase LabadfadfwoiejcÎncă nu există evaluări
- Amylase LabDocument2 paginiAmylase LabadfadfwoiejcÎncă nu există evaluări
- Amylase LabDocument2 paginiAmylase LabadfadfwoiejcÎncă nu există evaluări
- Enzyme Activity Factors and Plant Physiological ProcessesDocument4 paginiEnzyme Activity Factors and Plant Physiological ProcessesGeraldineMayÎncă nu există evaluări
- 1 V 2 NDocument20 pagini1 V 2 NGeraldineMayÎncă nu există evaluări
- Cheeseburger With Tater TotsDocument1 paginăCheeseburger With Tater TotsGeraldineMayÎncă nu există evaluări
- Achievements - History Church - First StageDocument9 paginiAchievements - History Church - First StageGeraldineMayÎncă nu există evaluări
- Science (Mains MCQ Part-2)Document24 paginiScience (Mains MCQ Part-2)Rajat KumarÎncă nu există evaluări
- Grade 8 Health 3rd Quarter ExamDocument2 paginiGrade 8 Health 3rd Quarter ExamHendrix Antonni AmanteÎncă nu există evaluări
- Arritmias Guia EscDocument36 paginiArritmias Guia EscKenny CantonÎncă nu există evaluări
- Concepts of Preload and AfterloadDocument2 paginiConcepts of Preload and AfterloadShar RiveraÎncă nu există evaluări
- Care Plan 5Document13 paginiCare Plan 5مالك مناصرةÎncă nu există evaluări
- Nonthrombogenic Roles of The Left Atrial AppendageDocument13 paginiNonthrombogenic Roles of The Left Atrial AppendageERIK EDUARDO BRICEÑO GÓMEZÎncă nu există evaluări
- An057 enDocument10 paginiAn057 enmehdi.chlif4374Încă nu există evaluări
- PHYSICAL Education Midterm Q1Document53 paginiPHYSICAL Education Midterm Q1Riki Leigh Palattao100% (1)
- 309 Lab ReportDocument8 pagini309 Lab ReportZorina Bi0% (1)
- 7th Grade Natural Science Reinforcement WorksheetDocument3 pagini7th Grade Natural Science Reinforcement WorksheetnataliaÎncă nu există evaluări
- How Animals Survive (Circulation and GasDocument24 paginiHow Animals Survive (Circulation and GasJohn Ted BañezÎncă nu există evaluări
- Comprehensive revision test - Basics 1-5Document16 paginiComprehensive revision test - Basics 1-5drpnnreddyÎncă nu există evaluări
- FocusCardiacUltra Dec2019 PDFDocument4 paginiFocusCardiacUltra Dec2019 PDFMeta Restu SynthanaÎncă nu există evaluări
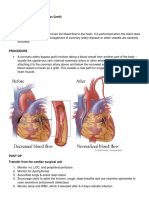
- CABG Surgery ExplainedDocument2 paginiCABG Surgery ExplainedebiiÎncă nu există evaluări
- 2005 10 24 Review AnswersDocument30 pagini2005 10 24 Review Answersaip1000Încă nu există evaluări
- BiomdcDocument11 paginiBiomdcNurSyafiqahAbdulHalimÎncă nu există evaluări
- ANPH111 Cardiovascular System OverviewDocument7 paginiANPH111 Cardiovascular System OverviewUwen NalpÎncă nu există evaluări
- Maintaining Exercise Progress in the Maintenance StageDocument7 paginiMaintaining Exercise Progress in the Maintenance StageSheena Mae PoÎncă nu există evaluări
- Uwrt 1103 - Family Tree EssayDocument6 paginiUwrt 1103 - Family Tree Essayapi-301005929Încă nu există evaluări
- The Artificial Heart: A Design Example: BIOE 1000 October 18, 2001Document17 paginiThe Artificial Heart: A Design Example: BIOE 1000 October 18, 2001Paspulati Leelaram100% (1)
- Proper Vital Signs MonitoringDocument7 paginiProper Vital Signs MonitoringCristoper BodionganÎncă nu există evaluări
- Cardiovascular Perfusion ThesisDocument4 paginiCardiovascular Perfusion Thesisamandagraytulsa100% (2)
- Liver, Biliary Tract and Pancreas Conditions ClassificationDocument69 paginiLiver, Biliary Tract and Pancreas Conditions Classificationdokter jaga hasna medikaÎncă nu există evaluări