S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Valmet DNA Machine Monitoring: Power Plants Erkki JaatinenDocument65 paginiValmet DNA Machine Monitoring: Power Plants Erkki JaatinenSabariyantoÎncă nu există evaluări
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- 05 PumpsDocument80 pagini05 PumpsRoberto O'ConnorÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Flow Sheet Post MC O2 Wash Press 1552018Document1 paginăFlow Sheet Post MC O2 Wash Press 1552018SabariyantoÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- SGA Reduce Active Chlorine ConsumptionDocument8 paginiSGA Reduce Active Chlorine ConsumptionSabariyantoÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- PRW Pulp Mill: Rusty IdentificationDocument12 paginiPRW Pulp Mill: Rusty IdentificationSabariyantoÎncă nu există evaluări
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- USttcDocument7 paginiUSttcSabariyantoÎncă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Silabus Training SGA WaterDocument3 paginiSilabus Training SGA WaterSabariyantoÎncă nu există evaluări
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- PRW Mill: Batch Cooking Fiberline 1Document9 paginiPRW Mill: Batch Cooking Fiberline 1SabariyantoÎncă nu există evaluări
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Anggit Dwi - 121160165 - Msds Sulfur DioxideDocument9 paginiAnggit Dwi - 121160165 - Msds Sulfur DioxideAnggit Dwi WÎncă nu există evaluări
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- GiDocument5 paginiGiSabariyantoÎncă nu există evaluări
- Transformer 3 PDFDocument80 paginiTransformer 3 PDFErix KhediraÎncă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- B SafeunitDocument4 paginiB SafeunitSabariyantoÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Wr0011black Liquor PDFDocument10 paginiWr0011black Liquor PDFSabariyantoÎncă nu există evaluări
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Material Safety Data Sheet (MSDS) Sulphur Dioxide (Please Ensure That This MSDS Is Received by The Appropriate Person)Document3 paginiMaterial Safety Data Sheet (MSDS) Sulphur Dioxide (Please Ensure That This MSDS Is Received by The Appropriate Person)Jency ModiÎncă nu există evaluări
- YfyufDocument2 paginiYfyufSabariyantoÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Meeting All Your Tubing Needs: Boiler TubesDocument2 paginiMeeting All Your Tubing Needs: Boiler TubesSabariyantoÎncă nu există evaluări
- BL MsdsDocument8 paginiBL MsdsSabariyantoÎncă nu există evaluări
- Chlorine Dioxide Health and SafetyDocument4 paginiChlorine Dioxide Health and SafetySabariyantoÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- Chlorine Dioxide PDFDocument4 paginiChlorine Dioxide PDFSudhakar Rao100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Mill Wide OptimizationDocument14 paginiMill Wide OptimizationSabariyantoÎncă nu există evaluări
- 1Document2 pagini1SabariyantoÎncă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Effective Separation of Foreign Material: Valmet Blow Line SeparatorDocument1 paginăEffective Separation of Foreign Material: Valmet Blow Line SeparatorSabariyantoÎncă nu există evaluări
- Ashleach: Increased Recovery Boiler AvailabilityDocument4 paginiAshleach: Increased Recovery Boiler AvailabilitySabariyantoÎncă nu există evaluări
- Bypass BS Filtrat 1 To Buffer Tank-Spill TankDocument2 paginiBypass BS Filtrat 1 To Buffer Tank-Spill TankSabariyantoÎncă nu există evaluări
- Inlet Pipe HurDocument4 paginiInlet Pipe HurSabariyantoÎncă nu există evaluări
- Fa Brochure PPC IkpDocument1 paginăFa Brochure PPC IkpSabariyantoÎncă nu există evaluări
- Inlet Pipe HurDocument4 paginiInlet Pipe HurSabariyantoÎncă nu există evaluări
- PROPOSAL 20180 IrirDocument70 paginiPROPOSAL 20180 IrirSabariyantoÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Inlet Pipe HurDocument4 paginiInlet Pipe HurSabariyantoÎncă nu există evaluări
- Standard Shaft: NOMINAL TOLERANCE UNIT Um (1 Um 0,001 MM)Document2 paginiStandard Shaft: NOMINAL TOLERANCE UNIT Um (1 Um 0,001 MM)SabariyantoÎncă nu există evaluări
- Chronic Fatigue Syndrome Caused by Electromagnetic Fields (EMF) ?Document6 paginiChronic Fatigue Syndrome Caused by Electromagnetic Fields (EMF) ?SwissTeslaÎncă nu există evaluări
- Immunopharmacology - IntroductionDocument51 paginiImmunopharmacology - IntroductionIsrat Jahan Surovy 2225400672Încă nu există evaluări
- Pages From 1894 Carey The Biochemic System of Medicine-Part 1of 9Document51 paginiPages From 1894 Carey The Biochemic System of Medicine-Part 1of 9Shiva Maru100% (3)
- K-2-Kuda - Digestive MonogastricDocument13 paginiK-2-Kuda - Digestive Monogastricsandy rianÎncă nu există evaluări
- Manual de RCP - NEONATAL - 7°ED PDFDocument328 paginiManual de RCP - NEONATAL - 7°ED PDFNatanael Librado97% (29)
- In Partial Fulfillment of The Final Requirements in Advanced Health Assessment MSN 205Document13 paginiIn Partial Fulfillment of The Final Requirements in Advanced Health Assessment MSN 205Gummie Akalal SugalaÎncă nu există evaluări
- Azzam Management of PreeclampsiaDocument36 paginiAzzam Management of PreeclampsiaHezal Zamzuri HudayaÎncă nu există evaluări
- Health in The Late Pre-Colonial and Early Colonial Period in The PhilippinesDocument237 paginiHealth in The Late Pre-Colonial and Early Colonial Period in The PhilippinesCurtiusRansÎncă nu există evaluări
- Should CO2 Be A Critical Process Parameter WhiteDocument4 paginiShould CO2 Be A Critical Process Parameter WhitesalemÎncă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Disease of Blood VesselsDocument5 paginiDisease of Blood VesselsKeshant SamarooÎncă nu există evaluări
- Management of Osteoporosis in CKD: Pascale Khairallah and Thomas L. NickolasDocument8 paginiManagement of Osteoporosis in CKD: Pascale Khairallah and Thomas L. NickolasSlr RandiÎncă nu există evaluări
- Spirometry in PCDocument12 paginiSpirometry in PCDanielleÎncă nu există evaluări
- OB Nursing-Q&ADocument46 paginiOB Nursing-Q&ABelle Moltio-BarcelonaÎncă nu există evaluări
- Cabg 130102113345 Phpapp01Document29 paginiCabg 130102113345 Phpapp01md.dascalescu2486Încă nu există evaluări
- Chapter 10 - Respiratory System - 2020 - The Zebrafish in Biomedical ResearchDocument5 paginiChapter 10 - Respiratory System - 2020 - The Zebrafish in Biomedical ResearchNicolas BaronÎncă nu există evaluări
- AGE NCPDocument4 paginiAGE NCPXane Tañada DiazÎncă nu există evaluări
- Pathology, Lecture 6 (Lecture Notes)Document18 paginiPathology, Lecture 6 (Lecture Notes)Ali Al-Qudsi100% (1)
- Assessment of The Critically Ill PatientsDocument30 paginiAssessment of The Critically Ill Patientsefancoolhand09Încă nu există evaluări
- 0610 s12 QP 11Document16 pagini0610 s12 QP 11Choudhry WalidÎncă nu există evaluări
- Comlex High YieldDocument4 paginiComlex High Yieldjoey plouffeÎncă nu există evaluări
- Revision Practice Questions - Cells & TransportDocument11 paginiRevision Practice Questions - Cells & TransportMarina BrazendaleÎncă nu există evaluări
- Goodnight Beautiful-Dorothy KoomsonDocument13 paginiGoodnight Beautiful-Dorothy KoomsondarijarasetaÎncă nu există evaluări
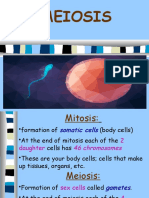
- MeiosisDocument39 paginiMeiosisGrachiella JanenÎncă nu există evaluări
- My Journey With Mum & MDSDocument54 paginiMy Journey With Mum & MDSIlene OngÎncă nu există evaluări
- CancerDocument75 paginiCancerJohn-john AngloÎncă nu există evaluări
- Respiratory Failure (Aan) PDFDocument19 paginiRespiratory Failure (Aan) PDFYudionoÎncă nu există evaluări
- INTEGUMENTARY SYSTEM - Review MaterialDocument11 paginiINTEGUMENTARY SYSTEM - Review MaterialSALGIE SERNALÎncă nu există evaluări
- Errors of RefractionDocument27 paginiErrors of RefractionAminaÎncă nu există evaluări
- Imb Be: Your Guide To Liv Ngina Ver Table WonderlandDocument17 paginiImb Be: Your Guide To Liv Ngina Ver Table WonderlandIlona HezingerÎncă nu există evaluări
- Cell Modification ModuleDocument30 paginiCell Modification ModuleDuane100% (1)
- The Obesity Code: Unlocking the Secrets of Weight LossDe la EverandThe Obesity Code: Unlocking the Secrets of Weight LossEvaluare: 4 din 5 stele4/5 (6)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDe la EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityEvaluare: 4 din 5 stele4/5 (28)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDe la EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsÎncă nu există evaluări
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDe la EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeEvaluare: 2 din 5 stele2/5 (1)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDe la EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDEvaluare: 5 din 5 stele5/5 (2)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDe la EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionEvaluare: 4 din 5 stele4/5 (404)