S-ar putea să vă placă și
- Amazon Rainforest Deforestation in the Lungs of the EarthDe la EverandAmazon Rainforest Deforestation in the Lungs of the EarthÎncă nu există evaluări
- Amazon'sDocument8 paginiAmazon'sStephanie N'goranÎncă nu există evaluări
- Aragao - s41467 017 02771 yDocument12 paginiAragao - s41467 017 02771 yRuth Nuria Alexandra Holgado ChallcoÎncă nu există evaluări
- Sena 2012Document12 paginiSena 2012ruanÎncă nu există evaluări
- The Future Climate of Amazonia ReportDocument42 paginiThe Future Climate of Amazonia ReportAlina JalbaÎncă nu există evaluări
- Nobre2009 PDFDocument9 paginiNobre2009 PDFJuanito Sin TierraÎncă nu există evaluări
- Tobon Et Al., 2000Document18 paginiTobon Et Al., 2000LAURA CATALINA VARGAS LENISÎncă nu există evaluări
- Scientific FrameworkDocument17 paginiScientific Frameworkym.mir4iÎncă nu există evaluări
- A Scientific Framework To Save The Amazon Final DraftDocument13 paginiA Scientific Framework To Save The Amazon Final DraftLuis Bedoya JaramilloÎncă nu există evaluări
- Tree-Ring Reconstructed Rainfall Over The Southern Amazon Basin: Southern Amazon RainfallDocument10 paginiTree-Ring Reconstructed Rainfall Over The Southern Amazon Basin: Southern Amazon Rainfalltaucci123Încă nu există evaluări
- Plant Communities of Western Amazonia: Bot. Rev. (2009) 75:271 - 291 DOI 10.1007/s12229-009-9032-1Document21 paginiPlant Communities of Western Amazonia: Bot. Rev. (2009) 75:271 - 291 DOI 10.1007/s12229-009-9032-1sandra hernandezÎncă nu există evaluări
- Gallery Springer 2014 PDFDocument23 paginiGallery Springer 2014 PDFarreeÎncă nu există evaluări
- ClimateDocument25 paginiClimateCintia DiasÎncă nu există evaluări
- Environmental DegradationDocument17 paginiEnvironmental DegradationmachariaÎncă nu există evaluări
- 13 - Variabilidade Mangue (Grupo 1)Document17 pagini13 - Variabilidade Mangue (Grupo 1)Letícia PortugalÎncă nu există evaluări
- Aquatic Ecosystem Health & ManagementDocument22 paginiAquatic Ecosystem Health & ManagementAlessandra MendonçaÎncă nu există evaluări
- Ampuero Et Al 2020 JGRDocument16 paginiAmpuero Et Al 2020 JGRLenin SucaÎncă nu există evaluări
- Variability of Mangrove Brazilian Coast Ecosystems Along The - Schaeffer-Novelli Et Al. (1990a)Document15 paginiVariability of Mangrove Brazilian Coast Ecosystems Along The - Schaeffer-Novelli Et Al. (1990a)Ricardo R de CarvalhoÎncă nu există evaluări
- Journal Pre-Proof: Science of The Total EnvironmentDocument26 paginiJournal Pre-Proof: Science of The Total EnvironmentYovana Avril Valles AraujoÎncă nu există evaluări
- Killeen&Solorzano RSTB2007Document8 paginiKilleen&Solorzano RSTB2007Tim KilleenÎncă nu există evaluări
- Whitmore 1998Document10 paginiWhitmore 1998Khalid HafazallahÎncă nu există evaluări
- Critical Transitions in The Amazon Forest System: AnalysisDocument25 paginiCritical Transitions in The Amazon Forest System: AnalysisdedizituÎncă nu există evaluări
- ReforestaciónDocument19 paginiReforestaciónDerly GómezÎncă nu există evaluări
- United Nations Sustainable Development Goal 13 - Climate ActionDocument5 paginiUnited Nations Sustainable Development Goal 13 - Climate ActionJieshun WangÎncă nu există evaluări
- Slash and Burn and Wetland Agriculture CLM 2013Document15 paginiSlash and Burn and Wetland Agriculture CLM 2013api-218069865Încă nu există evaluări
- Alves 2020Document14 paginiAlves 2020Matheus Bonifácio PamplonaÎncă nu există evaluări
- Climate Change in Brazil: Perspective On The Biogeochemistry of Inland WatersDocument14 paginiClimate Change in Brazil: Perspective On The Biogeochemistry of Inland WatersMatheus Bonifácio PamplonaÎncă nu există evaluări
- Chaves Et Al-2008-Hydrological ProcessesDocument11 paginiChaves Et Al-2008-Hydrological Processesroneyazevedo15Încă nu există evaluări
- Hoyos Et Al 2021 GEB ForestSavannaSouthAmDocument15 paginiHoyos Et Al 2021 GEB ForestSavannaSouthAmEstefaníaCeballosÎncă nu există evaluări
- Effects of Reduced Water Flow in A Riparian Forest Community: A Conservation ApproachDocument13 paginiEffects of Reduced Water Flow in A Riparian Forest Community: A Conservation ApproachAlejandro GilÎncă nu există evaluări
- CS#2 Human Impact On EcosystemsDocument1 paginăCS#2 Human Impact On EcosystemsJohn Andrae MangloÎncă nu există evaluări
- GlaciarDocument12 paginiGlaciarGian Carlo SuárezÎncă nu există evaluări
- Amazon Rainforest EssayDocument10 paginiAmazon Rainforest EssayJuan Esteban Castaño ArbelaezÎncă nu există evaluări
- Impacts of Hydropower and Climate Change On Drivers of Ecological Productivity of Southeast Asia's Most Important WetlandDocument12 paginiImpacts of Hydropower and Climate Change On Drivers of Ecological Productivity of Southeast Asia's Most Important WetlandNicholas de LeónÎncă nu există evaluări
- Growth and Water Balance of Eucalyptus Grandis Hybrid Plantations in Brazil During A Rotation For Pulp ProductionDocument6 paginiGrowth and Water Balance of Eucalyptus Grandis Hybrid Plantations in Brazil During A Rotation For Pulp ProductionGustavo Facincani DouradoÎncă nu există evaluări
- JGR Atmospheres - 2020 - Ampuero - The Forest Effects On The Isotopic Composition of Rainfall in The Northwestern AmazonDocument16 paginiJGR Atmospheres - 2020 - Ampuero - The Forest Effects On The Isotopic Composition of Rainfall in The Northwestern AmazonEDGAR MARCO FERANDEZ MEGOÎncă nu există evaluări
- Campo Lewis 2000Document12 paginiCampo Lewis 2000Windy HernandezÎncă nu există evaluări
- BosqueCuencayAgua PDFDocument15 paginiBosqueCuencayAgua PDFPedro Garrido VásquezÎncă nu există evaluări
- Certificate: Amazon RainforestDocument38 paginiCertificate: Amazon RainforestSuhas SurywanshiÎncă nu există evaluări
- Climate Change - News - Study Finds Severe Climate Jeopardizing Amazon ForestDocument2 paginiClimate Change - News - Study Finds Severe Climate Jeopardizing Amazon ForestRafael Gomes PereiraÎncă nu există evaluări
- Affonso Et Al, 2011Document14 paginiAffonso Et Al, 2011Lucas PaschoalÎncă nu există evaluări
- Joc 1475Document15 paginiJoc 1475Cleber AssisÎncă nu există evaluări
- 01 BagusDocument23 pagini01 Bagusfaiz ismaÎncă nu există evaluări
- Hutyra CO2 H2O Flux Tapajos 2007Document16 paginiHutyra CO2 H2O Flux Tapajos 2007Lucas BauerÎncă nu există evaluări
- Global Change Biology - 2006 - SCH NGART - Climate Growth Relationships of Tropical Tree Species in West Africa and TheirDocument12 paginiGlobal Change Biology - 2006 - SCH NGART - Climate Growth Relationships of Tropical Tree Species in West Africa and Theirbabie amadoÎncă nu există evaluări
- JOHNSON Et Al 2003 River and Stream EcosystemsDocument9 paginiJOHNSON Et Al 2003 River and Stream EcosystemsgulyÎncă nu există evaluări
- A Land Cover Map of South AmericaDocument14 paginiA Land Cover Map of South AmericaWander MacedoÎncă nu există evaluări
- CC AmphibsDocument13 paginiCC Amphibsjason manalotoÎncă nu există evaluări
- Ronchail Et Al-2002-International Journal of ClimatologyDocument24 paginiRonchail Et Al-2002-International Journal of ClimatologyAyrton OliveiraÎncă nu există evaluări
- The Vulnerability of Amazon Freshwater EcosystemsDocument13 paginiThe Vulnerability of Amazon Freshwater EcosystemsmfaunaÎncă nu există evaluări
- Paleoclimatic and Paleoenvironmental Changes in Amazonian Lowlands Over The LastDocument39 paginiPaleoclimatic and Paleoenvironmental Changes in Amazonian Lowlands Over The LastMIGUEL ANTONIO PALACIOS ARCEÎncă nu există evaluări
- Restrepo-Coupe Photosynthesis Seasonality Amazonia Brazil Flux Network 2013Document17 paginiRestrepo-Coupe Photosynthesis Seasonality Amazonia Brazil Flux Network 2013Lucas BauerÎncă nu există evaluări
- Costa, Botta, Cardille - Effects of Large-Scale Changes in Land Cover On The Discharge of The Tocantins River, Southeastern Amazonia - 2 PDFDocument12 paginiCosta, Botta, Cardille - Effects of Large-Scale Changes in Land Cover On The Discharge of The Tocantins River, Southeastern Amazonia - 2 PDFCleber AssisÎncă nu există evaluări
- Strengthening of Amazonian PrecipitationDocument12 paginiStrengthening of Amazonian PrecipitationblopamÎncă nu există evaluări
- Water and ForestsDocument15 paginiWater and Forestsjoeynarciso17Încă nu există evaluări
- Impact of Land Cover Change On Climate and El Niño in AustraliaDocument8 paginiImpact of Land Cover Change On Climate and El Niño in AustraliaDr Ravinesh C DeoÎncă nu există evaluări
- Franco 2014Document20 paginiFranco 2014Ariadne Cristina De AntonioÎncă nu există evaluări
- Zemp Et Al-2017-Geophysical Research LettersDocument9 paginiZemp Et Al-2017-Geophysical Research Lettersvass8464Încă nu există evaluări
- LessonDocument1 paginăLessonNeña Bautista PagalÎncă nu există evaluări
- Fagre Peterson 3-xDocument8 paginiFagre Peterson 3-xliboaninoÎncă nu există evaluări
- 934 019 002 PDFDocument187 pagini934 019 002 PDFNoCWÎncă nu există evaluări
- Moulinex Catalogue 2016 EnglishDocument25 paginiMoulinex Catalogue 2016 EnglishNoCWÎncă nu există evaluări
- Scyan X7 User GuideDocument9 paginiScyan X7 User GuideNoCWÎncă nu există evaluări
- H Frame Folding Tabletop Easel PlanDocument25 paginiH Frame Folding Tabletop Easel PlanFiorenzoAlbatrosÎncă nu există evaluări
- Intermec Program PrintDocument385 paginiIntermec Program PrintbravemanvvnÎncă nu există evaluări
- Moulinex BreadMaker Recipe BookDocument12 paginiMoulinex BreadMaker Recipe BookKar Gayee0% (1)
- Cloud Licensing OracleDocument2 paginiCloud Licensing OracleXan OrnelasÎncă nu există evaluări
- HelpfulnessDocument10 paginiHelpfulnessNoCWÎncă nu există evaluări
- IpeirotisDocument4 paginiIpeirotisNoCWÎncă nu există evaluări
- Informe Amazonia Viva 1990 2010Document60 paginiInforme Amazonia Viva 1990 2010Agustin Mamani MaytaÎncă nu există evaluări
- Lessons From The Amazon Picking Challenge: Four Aspects of Building Robotic SystemsDocument10 paginiLessons From The Amazon Picking Challenge: Four Aspects of Building Robotic SystemsNoCWÎncă nu există evaluări
- Language Demographics of Mechanical TurkDocument13 paginiLanguage Demographics of Mechanical TurkNoCWÎncă nu există evaluări
- Amazon So YaDocument64 paginiAmazon So YaNoCWÎncă nu există evaluări
- Informe Amazonia Viva 1990 2010Document60 paginiInforme Amazonia Viva 1990 2010Agustin Mamani MaytaÎncă nu există evaluări
- Technical Report TR-08-07: An Evaluation of Amazon's Grid Computing Services: EC2, S3 and SQSDocument15 paginiTechnical Report TR-08-07: An Evaluation of Amazon's Grid Computing Services: EC2, S3 and SQSNoCWÎncă nu există evaluări
- Amazon So YaDocument64 paginiAmazon So YaNoCWÎncă nu există evaluări
- Cloud Licensing OracleDocument2 paginiCloud Licensing OracleXan OrnelasÎncă nu există evaluări
- #11a What Makes A Helpful Online Review A Study of Customer Reviews OnDocument17 pagini#11a What Makes A Helpful Online Review A Study of Customer Reviews OnDicky PradanaÎncă nu există evaluări
- Amazon OrderDocument23 paginiAmazon OrderNoCWÎncă nu există evaluări
- 1853 PalmTrees S713 PDFDocument248 pagini1853 PalmTrees S713 PDFNoCWÎncă nu există evaluări
- Thoughts Ereaders PDFDocument8 paginiThoughts Ereaders PDFNoCWÎncă nu există evaluări
- The Effectiveness of Climate Finance: A Review of The Amazon FundDocument38 paginiThe Effectiveness of Climate Finance: A Review of The Amazon FundNoCWÎncă nu există evaluări
- R42629Document13 paginiR42629NoCWÎncă nu există evaluări
- Lessons From The Amazon Picking Challenge: Four Aspects of Building Robotic SystemsDocument10 paginiLessons From The Amazon Picking Challenge: Four Aspects of Building Robotic SystemsNoCWÎncă nu există evaluări
- Scientific Computing Using Virtual High-Performance Computing PDFDocument10 paginiScientific Computing Using Virtual High-Performance Computing PDFNoCWÎncă nu există evaluări
- Hamburger Connection Fuels Amazon Destruction1 PDFDocument10 paginiHamburger Connection Fuels Amazon Destruction1 PDFNoCWÎncă nu există evaluări
- Amazon 10 K 2015Document86 paginiAmazon 10 K 2015NoCWÎncă nu există evaluări
- Timothy Leung Introduction To AlibabaDocument15 paginiTimothy Leung Introduction To AlibabaMubashir Ali KhanÎncă nu există evaluări
- Amazon FinalDocument45 paginiAmazon FinalNoCWÎncă nu există evaluări
- BdeM Taller No. 2Document2 paginiBdeM Taller No. 2Luisfer PriceÎncă nu există evaluări
- Transcritical R744 (CO2) Heat Pumps: October 2007Document64 paginiTranscritical R744 (CO2) Heat Pumps: October 2007حسن عرقسوسيÎncă nu există evaluări
- Produção de OxigênioDocument30 paginiProdução de OxigênioLeonardo CarneiroÎncă nu există evaluări
- Coal Chapter 3Document24 paginiCoal Chapter 3Ihwan Limanto100% (1)
- EssayDocument3 paginiEssayapi-438118584Încă nu există evaluări
- Ident-No: ARADUR HY 951: Safety Data SheetDocument7 paginiIdent-No: ARADUR HY 951: Safety Data SheetPaulÎncă nu există evaluări
- SCOPE 21 - The Major Biogeochemical Cycles and Their InteractionsDocument205 paginiSCOPE 21 - The Major Biogeochemical Cycles and Their InteractionsAlina JalbaÎncă nu există evaluări
- Apocalypse Humans ProjectDocument18 paginiApocalypse Humans ProjectaubweeÎncă nu există evaluări
- SAPS - Photosynthesis With Algal Balls - Teachers NotesDocument10 paginiSAPS - Photosynthesis With Algal Balls - Teachers NotesHisham El KanayatiÎncă nu există evaluări
- Mcqs On EnvironmentDocument17 paginiMcqs On EnvironmentMasroor AhmadÎncă nu există evaluări
- Practice KEY - Stoichiometry PAP 2019-2020-4Document16 paginiPractice KEY - Stoichiometry PAP 2019-2020-4toxxic21Încă nu există evaluări
- Carbon Dioxide Utilization in Ready-Mixed Concrete ProductionDocument24 paginiCarbon Dioxide Utilization in Ready-Mixed Concrete Productionjack21ab100% (1)
- Chapter 19 Equations of Change For Multicomponent SystemsDocument37 paginiChapter 19 Equations of Change For Multicomponent SystemsJohnÎncă nu există evaluări
- Theory of CombustionDocument19 paginiTheory of CombustionDevendra Singh DhakreyÎncă nu există evaluări
- Msds Sigma Phenguard 930 (Sigma Phenguard Primer) BaseDocument7 paginiMsds Sigma Phenguard 930 (Sigma Phenguard Primer) BasekumarÎncă nu există evaluări
- Dcem2100 - Tuv - Ops (0001)Document66 paginiDcem2100 - Tuv - Ops (0001)Quan Nguyen VanÎncă nu există evaluări
- Mastery Assignment 1 Lab Report 1Document7 paginiMastery Assignment 1 Lab Report 1Peter MungaiÎncă nu există evaluări
- CO2 Storage Tank Product Manual WsDocument34 paginiCO2 Storage Tank Product Manual WsGalang Hanif AbdulahÎncă nu există evaluări
- 1.5 HPHT Testing PDFDocument6 pagini1.5 HPHT Testing PDFHATEMÎncă nu există evaluări
- Greenhouse Climate Control An Integrated Approach PDFDocument293 paginiGreenhouse Climate Control An Integrated Approach PDFraj100% (2)
- PPTPV (SDS) en - Marabu ThinnerDocument8 paginiPPTPV (SDS) en - Marabu ThinnerNisma NilamsariÎncă nu există evaluări
- Oil and Fat Technology Lectures IIDocument24 paginiOil and Fat Technology Lectures IIaulger100% (1)
- Class 7 Science Exam Paper March 2021-22Document9 paginiClass 7 Science Exam Paper March 2021-22Riffat AamerÎncă nu există evaluări
- The Impact of CO2 On Childrens Learning WebDocument12 paginiThe Impact of CO2 On Childrens Learning WebKaterinaÎncă nu există evaluări
- Modeling CO2 Storage in Aquifers With A Fully-CoupDocument17 paginiModeling CO2 Storage in Aquifers With A Fully-CoupFatimah Az ZahraÎncă nu există evaluări
- CEMS RATA ProtocolDocument101 paginiCEMS RATA ProtocolALI RAZAÎncă nu există evaluări
- F1 资料大全Document18 paginiF1 资料大全Sugar LeeÎncă nu există evaluări
- 1Document9 pagini1Silambarasan SivalingamÎncă nu există evaluări
- Linde N2 EOR Case Study CantarellDocument26 paginiLinde N2 EOR Case Study CantarellEliza NunesÎncă nu există evaluări
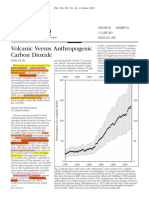
- Volcanic Versus Anthropogenic Carbon Dioxide (2011 EOS) PDFDocument3 paginiVolcanic Versus Anthropogenic Carbon Dioxide (2011 EOS) PDFsuperearthÎncă nu există evaluări
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessDe la EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessÎncă nu există evaluări
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincDe la EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincEvaluare: 3.5 din 5 stele3.5/5 (137)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingDe la EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingEvaluare: 3.5 din 5 stele3.5/5 (33)
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseDe la EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseEvaluare: 3.5 din 5 stele3.5/5 (69)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingDe la EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingEvaluare: 5 din 5 stele5/5 (5)
- Lessons for Survival: Mothering Against “the Apocalypse”De la EverandLessons for Survival: Mothering Against “the Apocalypse”Evaluare: 5 din 5 stele5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (811)
- The Storm of the Century: Tragedy, Heroism, Survival, and the Epic True Story of America's Deadliest Natural DisasterDe la EverandThe Storm of the Century: Tragedy, Heroism, Survival, and the Epic True Story of America's Deadliest Natural DisasterÎncă nu există evaluări
- Water: The Epic Struggle for Wealth, Power, and CivilizationDe la EverandWater: The Epic Struggle for Wealth, Power, and CivilizationEvaluare: 3.5 din 5 stele3.5/5 (37)
- Fire Season: Field Notes from a Wilderness LookoutDe la EverandFire Season: Field Notes from a Wilderness LookoutEvaluare: 4 din 5 stele4/5 (142)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDe la EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldEvaluare: 4.5 din 5 stele4.5/5 (18)
- World of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsDe la EverandWorld of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsEvaluare: 4 din 5 stele4/5 (223)
- Water to the Angels: William Mulholland, His Monumental Aqueduct, and the Rise of Los AngelesDe la EverandWater to the Angels: William Mulholland, His Monumental Aqueduct, and the Rise of Los AngelesEvaluare: 4 din 5 stele4/5 (21)
- Chesapeake Requiem: A Year with the Watermen of Vanishing Tangier IslandDe la EverandChesapeake Requiem: A Year with the Watermen of Vanishing Tangier IslandEvaluare: 4 din 5 stele4/5 (38)
- Smokejumper: A Memoir by One of America's Most Select Airborne FirefightersDe la EverandSmokejumper: A Memoir by One of America's Most Select Airborne FirefightersÎncă nu există evaluări
- The Other End of the Leash: Why We Do What We Do Around DogsDe la EverandThe Other End of the Leash: Why We Do What We Do Around DogsEvaluare: 5 din 5 stele5/5 (65)
- Come Back, Como: Winning the Heart of a Reluctant DogDe la EverandCome Back, Como: Winning the Heart of a Reluctant DogEvaluare: 3.5 din 5 stele3.5/5 (10)
- When You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsDe la EverandWhen You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsEvaluare: 3.5 din 5 stele3.5/5 (13)