S-ar putea să vă placă și
- ARTÍCULO 2. Lancet BiasL3Document5 paginiARTÍCULO 2. Lancet BiasL3Gabriela Zavaleta CamachoÎncă nu există evaluări
- ARTÍCULO 3. Observational Studies IFD L1 PDFDocument7 paginiARTÍCULO 3. Observational Studies IFD L1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 3-Pericontusional Brain Tissue Exhibits Persistent Elevation Of...Document8 pagini3-Pericontusional Brain Tissue Exhibits Persistent Elevation Of...Gabriela Zavaleta CamachoÎncă nu există evaluări
- 5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Document27 pagini5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Gabriela Zavaleta CamachoÎncă nu există evaluări
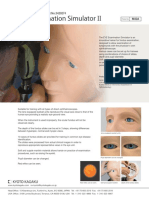
- Kyoto Kagaku - Eye Examination SimulatorDocument2 paginiKyoto Kagaku - Eye Examination SimulatorGabriela Zavaleta Camacho100% (1)
- 6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFDocument14 pagini6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- TCDocument17 paginiTCGabriela Zavaleta CamachoÎncă nu există evaluări
- Tool phq9 PDFDocument3 paginiTool phq9 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- BrainDocument51 paginiBrainAnonymous gMLTpER9IUÎncă nu există evaluări
- Kyoto Kagaku - Ear Examination SimulatorDocument2 paginiKyoto Kagaku - Ear Examination SimulatorGabriela Zavaleta CamachoÎncă nu există evaluări
- 6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFDocument14 pagini6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Document27 pagini5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Gabriela Zavaleta CamachoÎncă nu există evaluări
- 10-Lactate Brain Fuel in Human Traumatic Brain Injury. 17Document52 pagini10-Lactate Brain Fuel in Human Traumatic Brain Injury. 17Gabriela Zavaleta CamachoÎncă nu există evaluări
- 9-Lactate Uptake by The Injured Human Brain-18Document7 pagini9-Lactate Uptake by The Injured Human Brain-18Gabriela Zavaleta CamachoÎncă nu există evaluări
- 12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Document12 pagini12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Gabriela Zavaleta CamachoÎncă nu există evaluări
- 12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Document12 pagini12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Gabriela Zavaleta CamachoÎncă nu există evaluări
- 14-Current Trends in Lactate Metabolism Introduction.11Document2 pagini14-Current Trends in Lactate Metabolism Introduction.11Gabriela Zavaleta CamachoÎncă nu există evaluări
- 11 Glycolysisandthesignificanceoflactateintraumaticbraininjury 8Document15 pagini11 Glycolysisandthesignificanceoflactateintraumaticbraininjury 8Gabriela Zavaleta CamachoÎncă nu există evaluări
- Cerebral Metabolism Following Traumatic Brain Injury: New Discoveries With Implications For TreatmentDocument13 paginiCerebral Metabolism Following Traumatic Brain Injury: New Discoveries With Implications For TreatmentGabriela Zavaleta CamachoÎncă nu există evaluări
- 16-Cerebral Lactate Metabolism After Traumatic Brain Injury-15Document7 pagini16-Cerebral Lactate Metabolism After Traumatic Brain Injury-15Gabriela Zavaleta CamachoÎncă nu există evaluări
- Tool phq9 PDFDocument3 paginiTool phq9 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 18-Cerebral Extracellular Chemistry and Outcome-9Document11 pagini18-Cerebral Extracellular Chemistry and Outcome-9Gabriela Zavaleta CamachoÎncă nu există evaluări
- 10 1001@jama 2016 0287Document10 pagini10 1001@jama 2016 0287ompardor7554Încă nu există evaluări
- 19-Base Deficit and Serum Lactate Concentration in Tec-11Document5 pagini19-Base Deficit and Serum Lactate Concentration in Tec-11Gabriela Zavaleta CamachoÎncă nu există evaluări
- Serum Lactate As A Predictor of Mortality in Patients With InfectionDocument8 paginiSerum Lactate As A Predictor of Mortality in Patients With InfectionGabriela Zavaleta CamachoÎncă nu există evaluări
- Guideline CA Cervix EscociaDocument77 paginiGuideline CA Cervix EscociaGabriela Zavaleta CamachoÎncă nu există evaluări
- Depression Brazilian AdolescentsDocument6 paginiDepression Brazilian AdolescentsGabriela Zavaleta CamachoÎncă nu există evaluări
- Current Management Hipertension PDFDocument11 paginiCurrent Management Hipertension PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- PDFDocument25 paginiPDFTruly GracevaÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Aiapget 2020 QPDocument29 paginiAiapget 2020 QPGanesh RadhakrishnanÎncă nu există evaluări
- The Ultimate Guide To Anxiety DisordersDocument66 paginiThe Ultimate Guide To Anxiety Disordersnajaxx100% (2)
- The Lord of Lost Heart PDFDocument44 paginiThe Lord of Lost Heart PDFCouteron LaurentÎncă nu există evaluări
- Paedo Don TicsDocument103 paginiPaedo Don TicsIana RusuÎncă nu există evaluări
- Challenges and Opportunities in Preventive and Social MedicineDocument3 paginiChallenges and Opportunities in Preventive and Social MedicineIJAR JOURNALÎncă nu există evaluări
- 1 s2.0 S2772569323000026 MainDocument8 pagini1 s2.0 S2772569323000026 MainAditya RizkyÎncă nu există evaluări
- Lab Diagnosis of Diarrheal DiseasesDocument37 paginiLab Diagnosis of Diarrheal DiseasesGIST (Gujarat Institute of Science & Technology)Încă nu există evaluări
- Rat ExperimentDocument7 paginiRat ExperimentKavithaa Sengalrayan0% (1)
- Trends, issues, and magnitude of psychiatric nursingDocument4 paginiTrends, issues, and magnitude of psychiatric nursingsona0% (1)
- Periodontics PDFDocument195 paginiPeriodontics PDFدينا حسن نعمة100% (1)
- MRNA Vaccines - Day - 04.14.20 PDFDocument259 paginiMRNA Vaccines - Day - 04.14.20 PDFTammy G100% (1)
- Quality Assurance in Medical LaboratoriesDocument69 paginiQuality Assurance in Medical Laboratories"DocAxi" Maximo B Axibal Jr MD FPSP100% (1)
- Grade 4 WINS OrientationDocument2 paginiGrade 4 WINS OrientationChristine FranciscoÎncă nu există evaluări
- Lean GuideDocument155 paginiLean GuideSujoy Kar100% (1)
- Penicillin: Weird RPG Zine Issue 1 F A L L 2 0 1 9Document16 paginiPenicillin: Weird RPG Zine Issue 1 F A L L 2 0 1 9iÎncă nu există evaluări
- Mozart, Beethoven, Yesudas Effect Class 2Document12 paginiMozart, Beethoven, Yesudas Effect Class 2Dr Suvarna NalapatÎncă nu există evaluări
- STERILE PARENTERAL FORMULATIONSDocument85 paginiSTERILE PARENTERAL FORMULATIONSShalimashalu100% (1)
- Coping Strategies of Athletes for Competitive AnxietyDocument13 paginiCoping Strategies of Athletes for Competitive AnxietyHONEY JEAN ESPENORIOÎncă nu există evaluări
- Idnt Na MatrixDocument20 paginiIdnt Na MatrixicalÎncă nu există evaluări
- Fundamentals of EU Regulatory Affairs, Fifth Edition Comparative MatrixDocument42 paginiFundamentals of EU Regulatory Affairs, Fifth Edition Comparative Matrixasifmdzakaria57% (7)
- 004 General Consent English (Front)Document2 pagini004 General Consent English (Front)عبدالله الرعويÎncă nu există evaluări
- Tun Razak Exchange, Retail Plot 1: APPENDIX 5 - Incident Reporting ProcedureDocument5 paginiTun Razak Exchange, Retail Plot 1: APPENDIX 5 - Incident Reporting ProcedureMatthew Mohan PerumalÎncă nu există evaluări
- Warehouse Center Checklist 2Document5 paginiWarehouse Center Checklist 2Sankar ChinnathambiÎncă nu există evaluări
- Acy 087Document14 paginiAcy 087johnsmithdosÎncă nu există evaluări
- Bio SaftyDocument7 paginiBio Saftyapi-283956320Încă nu există evaluări
- LeaP PE G6 Week 1 Q3Document6 paginiLeaP PE G6 Week 1 Q3Rosemarie Mañabo RamirezÎncă nu există evaluări
- New Consensus On Non Cirrhotic Portal FibrosisDocument41 paginiNew Consensus On Non Cirrhotic Portal Fibrosisramesh5889Încă nu există evaluări
- Diuretics MOA, Examples, Effects, and Nursing ConsiderationsDocument1 paginăDiuretics MOA, Examples, Effects, and Nursing ConsiderationsGrace ButlerÎncă nu există evaluări
- Cipero & Guaracara River Water Quality Site VisitsDocument76 paginiCipero & Guaracara River Water Quality Site VisitsBrandon Atwell79% (29)
- Attachment Theory and FathersDocument16 paginiAttachment Theory and FathersGraceÎncă nu există evaluări