S-ar putea să vă placă și
- ARTÍCULO 2. Lancet BiasL3Document5 paginiARTÍCULO 2. Lancet BiasL3Gabriela Zavaleta CamachoÎncă nu există evaluări
- Kyoto Kagaku - Ear Examination SimulatorDocument2 paginiKyoto Kagaku - Ear Examination SimulatorGabriela Zavaleta CamachoÎncă nu există evaluări
- ARTÍCULO 3. Observational Studies IFD L1 PDFDocument7 paginiARTÍCULO 3. Observational Studies IFD L1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- Tool phq9 PDFDocument3 paginiTool phq9 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- TCDocument17 paginiTCGabriela Zavaleta CamachoÎncă nu există evaluări
- BrainDocument51 paginiBrainAnonymous gMLTpER9IUÎncă nu există evaluări
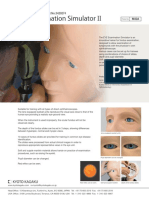
- Kyoto Kagaku - Eye Examination SimulatorDocument2 paginiKyoto Kagaku - Eye Examination SimulatorGabriela Zavaleta Camacho100% (1)
- 3-Pericontusional Brain Tissue Exhibits Persistent Elevation Of...Document8 pagini3-Pericontusional Brain Tissue Exhibits Persistent Elevation Of...Gabriela Zavaleta CamachoÎncă nu există evaluări
- 9-Lactate Uptake by The Injured Human Brain-18Document7 pagini9-Lactate Uptake by The Injured Human Brain-18Gabriela Zavaleta CamachoÎncă nu există evaluări
- 6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFDocument14 pagini6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFDocument14 pagini6-Moderate and Severe Traumatic Brain Injury in Adults - 1 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Document27 pagini5-Neuroenergetic Response To Prolonged Cerebral Glucose Depletion After Severe Brain Injury and The-20Gabriela Zavaleta CamachoÎncă nu există evaluări
- 11 Glycolysisandthesignificanceoflactateintraumaticbraininjury 8Document15 pagini11 Glycolysisandthesignificanceoflactateintraumaticbraininjury 8Gabriela Zavaleta CamachoÎncă nu există evaluări
- 10-Lactate Brain Fuel in Human Traumatic Brain Injury. 17Document52 pagini10-Lactate Brain Fuel in Human Traumatic Brain Injury. 17Gabriela Zavaleta CamachoÎncă nu există evaluări
- Cerebral Metabolism Following Traumatic Brain Injury: New Discoveries With Implications For TreatmentDocument13 paginiCerebral Metabolism Following Traumatic Brain Injury: New Discoveries With Implications For TreatmentGabriela Zavaleta CamachoÎncă nu există evaluări
- 17-Cerebral Extracellular Lactate Increase in Tec-19Document8 pagini17-Cerebral Extracellular Lactate Increase in Tec-19Gabriela Zavaleta CamachoÎncă nu există evaluări
- 14-Current Trends in Lactate Metabolism Introduction.11Document2 pagini14-Current Trends in Lactate Metabolism Introduction.11Gabriela Zavaleta CamachoÎncă nu există evaluări
- 12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Document12 pagini12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Gabriela Zavaleta CamachoÎncă nu există evaluări
- 12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Document12 pagini12-Energy Dysfunction As A Predictor of Outcome After Moderate Trauma-16Gabriela Zavaleta CamachoÎncă nu există evaluări
- 16-Cerebral Lactate Metabolism After Traumatic Brain Injury-15Document7 pagini16-Cerebral Lactate Metabolism After Traumatic Brain Injury-15Gabriela Zavaleta CamachoÎncă nu există evaluări
- Depression Brazilian AdolescentsDocument6 paginiDepression Brazilian AdolescentsGabriela Zavaleta CamachoÎncă nu există evaluări
- 18-Cerebral Extracellular Chemistry and Outcome-9Document11 pagini18-Cerebral Extracellular Chemistry and Outcome-9Gabriela Zavaleta CamachoÎncă nu există evaluări
- 19-Base Deficit and Serum Lactate Concentration in Tec-11Document5 pagini19-Base Deficit and Serum Lactate Concentration in Tec-11Gabriela Zavaleta CamachoÎncă nu există evaluări
- Serum Lactate As A Predictor of Mortality in Patients With InfectionDocument8 paginiSerum Lactate As A Predictor of Mortality in Patients With InfectionGabriela Zavaleta CamachoÎncă nu există evaluări
- Current Management Hipertension PDFDocument11 paginiCurrent Management Hipertension PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- Tool phq9 PDFDocument3 paginiTool phq9 PDFGabriela Zavaleta CamachoÎncă nu există evaluări
- 10 1001@jama 2016 0287Document10 pagini10 1001@jama 2016 0287ompardor7554Încă nu există evaluări
- PDFDocument25 paginiPDFTruly GracevaÎncă nu există evaluări
- Guideline CA Cervix EscociaDocument77 paginiGuideline CA Cervix EscociaGabriela Zavaleta CamachoÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Tefnol 1Document11 paginiTefnol 1Moustapha Salem MansourÎncă nu există evaluări
- As 400Document70 paginiAs 400Radhakrishnan KandhasamyÎncă nu există evaluări
- Introduction To GIS (Geographical Information System)Document20 paginiIntroduction To GIS (Geographical Information System)pwnjha100% (3)
- Zipato MQTTCloudDocument34 paginiZipato MQTTClouddensasÎncă nu există evaluări
- MagPi70 PDFDocument100 paginiMagPi70 PDFDaniel TuarezÎncă nu există evaluări
- Integrating Theory, Experiments, and FEA To Solve Challenging Nonlinear Mechanics ProblemsDocument17 paginiIntegrating Theory, Experiments, and FEA To Solve Challenging Nonlinear Mechanics ProblemsBodieTechÎncă nu există evaluări
- 02 Bateria 12V - 150AHDocument2 pagini02 Bateria 12V - 150AHRafael AlvarezÎncă nu există evaluări
- Table and Graph CriteriaDocument3 paginiTable and Graph Criteriaapi-271250844Încă nu există evaluări
- S3 3 TrigonometryDocument81 paginiS3 3 TrigonometryEugene ChoongÎncă nu există evaluări
- 20-F201007-SC-010 R0 - Design Calculation of Foam Tank and Fire Pump ShedDocument187 pagini20-F201007-SC-010 R0 - Design Calculation of Foam Tank and Fire Pump ShedSaravan KumarÎncă nu există evaluări
- Topsoe Ammonia 4 Start Up Worlds Largest Ammonia PlantDocument13 paginiTopsoe Ammonia 4 Start Up Worlds Largest Ammonia Plantpekanselandar0% (1)
- Random Numbers in PythonDocument3 paginiRandom Numbers in PythonShubham RawatÎncă nu există evaluări
- Shear Force & Bending Moment TestDocument11 paginiShear Force & Bending Moment TestKalaiArasanÎncă nu există evaluări
- MDM Heiana Nadia Hamzah: Prepared byDocument50 paginiMDM Heiana Nadia Hamzah: Prepared bySyarfa FurzanneÎncă nu există evaluări
- PIA B2 - Module 2 (PHYSICS) SubModule 2.2 (Mechanics) FinalDocument82 paginiPIA B2 - Module 2 (PHYSICS) SubModule 2.2 (Mechanics) Finalsamarrana1234679Încă nu există evaluări
- 4 A Short History of Islamic Science From Genesis To Decline ShortversionDocument24 pagini4 A Short History of Islamic Science From Genesis To Decline ShortversionKateryna KuslyvaÎncă nu există evaluări
- Fmaths 3RD Term YR11 PlanDocument28 paginiFmaths 3RD Term YR11 Planadegunloye temitopeÎncă nu există evaluări
- PHP Question AnswerDocument20 paginiPHP Question AnswerManish SharmaÎncă nu există evaluări
- Elliptical Head Design ToolDocument1 paginăElliptical Head Design ToolSudjono BroÎncă nu există evaluări
- A2 QuestionsDocument2 paginiA2 QuestionsJisu Elsa JacobÎncă nu există evaluări
- EE TermsDocument25 paginiEE TermsKerr AgotÎncă nu există evaluări
- 125 Tractor: (Specifications and Design Subject To Change Without Notice)Document5 pagini125 Tractor: (Specifications and Design Subject To Change Without Notice)Gary LarsonÎncă nu există evaluări
- Guest WiFi With MikroTik RoutersDocument1 paginăGuest WiFi With MikroTik Routersmahad3vaÎncă nu există evaluări
- (FreeCourseWeb - Com) 1493997599Document386 pagini(FreeCourseWeb - Com) 1493997599MuruganandamGanesanÎncă nu există evaluări
- Tài Liệu CAT Pallet Truck NPP20NDocument9 paginiTài Liệu CAT Pallet Truck NPP20NJONHHY NGUYEN DANGÎncă nu există evaluări
- Microstructure and Mechanical Properties of Borated Stainless Steel (304B) GTA and SMA WeldsDocument6 paginiMicrostructure and Mechanical Properties of Borated Stainless Steel (304B) GTA and SMA WeldsReza nugrahaÎncă nu există evaluări
- West Knits Book 3Document56 paginiWest Knits Book 3Alexandr Maxiuta100% (17)
- Seam Strength of Corrugated Plate With High Strength SteelDocument15 paginiSeam Strength of Corrugated Plate With High Strength SteelMariano SalcedoÎncă nu există evaluări
- C ValueDocument2 paginiC ValuePadmanabh Shetty0% (1)
- Es3Pt: User ManualDocument53 paginiEs3Pt: User ManualBITGEORGYÎncă nu există evaluări