S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Alternative Telomere RepaiirDocument16 paginiAlternative Telomere RepaiirMaithreyanNandhakumarÎncă nu există evaluări
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Extraintestinal Amebiasis': Scess Was PresentDocument3 paginiExtraintestinal Amebiasis': Scess Was PresentMaithreyanNandhakumarÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Present and Future Role of Microfluidics PDFDocument9 paginiThe Present and Future Role of Microfluidics PDFMaithreyanNandhakumarÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Aikat1979 PDFDocument5 paginiAikat1979 PDFMaithreyanNandhakumarÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- 4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerDocument18 pagini4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerMaithreyanNandhakumarÎncă nu există evaluări
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- ThreeChannel PDFDocument8 paginiThreeChannel PDFMaithreyanNandhakumarÎncă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Current Concept Same Bias IsDocument10 paginiCurrent Concept Same Bias IsMaithreyanNandhakumarÎncă nu există evaluări
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- 5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsDocument29 pagini5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsMaithreyanNandhakumarÎncă nu există evaluări
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Fate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyDocument8 paginiFate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyMaithreyanNandhakumarÎncă nu există evaluări
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- 3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningDocument11 pagini3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningMaithreyanNandhakumarÎncă nu există evaluări
- 2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfDocument25 pagini2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfMaithreyanNandhakumarÎncă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- 1microfluidic ModellingDocument22 pagini1microfluidic ModellingMaithreyanNandhakumarÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- ReferencesDocument4 paginiReferencesMaithreyanNandhakumarÎncă nu există evaluări
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Antifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2Document93 paginiAntifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2MaithreyanNandhakumarÎncă nu există evaluări
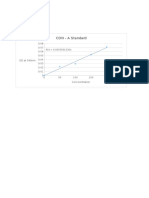
- CON - A Standard: OD at 540nmDocument10 paginiCON - A Standard: OD at 540nmMaithreyanNandhakumarÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Introduction To Flow Cytometry - A Learning GuideDocument54 paginiIntroduction To Flow Cytometry - A Learning GuideFrank FontaineÎncă nu există evaluări
- Focal Adhesion Dynamics in Cellular Function and DiseaseDocument19 paginiFocal Adhesion Dynamics in Cellular Function and DiseaseEly Huaman PorrasÎncă nu există evaluări
- Cell JunctionDocument63 paginiCell JunctionsphuyalÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- Harnessing Bene Fit From Targeting Tumor Associated Carbohydrate AntigensDocument9 paginiHarnessing Bene Fit From Targeting Tumor Associated Carbohydrate Antigensspin_echoÎncă nu există evaluări
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Gastrointestinal - Mucosal.repair - And.experimental - Therapeutics Ublog - TKDocument261 paginiGastrointestinal - Mucosal.repair - And.experimental - Therapeutics Ublog - TKpotaiskiÎncă nu există evaluări
- Translation and Its Role in CancerDocument709 paginiTranslation and Its Role in Cancerqasim_saeed_1Încă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- 4) 2023 Identification of A Novel Type of Focal Adhesion Remodelling Via FAKFRNK Replacement, and Its Contribution To Cancer ProgressionDocument15 pagini4) 2023 Identification of A Novel Type of Focal Adhesion Remodelling Via FAKFRNK Replacement, and Its Contribution To Cancer ProgressionFranc DechiaÎncă nu există evaluări
- Neuromuscular Disorders - A. Zaher (Intech, 2012) WW PDFDocument280 paginiNeuromuscular Disorders - A. Zaher (Intech, 2012) WW PDFDaianaÎncă nu există evaluări
- Pharmacological ResearchDocument12 paginiPharmacological ResearchJUAN DIEGO TRUJILLO ROJASÎncă nu există evaluări
- Strength Training in Search of Optimal Strategies To Maximize Neuromuscular Performance.Document13 paginiStrength Training in Search of Optimal Strategies To Maximize Neuromuscular Performance.Hugo TintiÎncă nu există evaluări
- The Avp3 Integrin "Vitronectin Receptor": Molecules in FocusDocument5 paginiThe Avp3 Integrin "Vitronectin Receptor": Molecules in FocusvadhaÎncă nu există evaluări
- Creating Avenues - Spreading Truth: A Research Awareness E-Journal by IPRHDocument52 paginiCreating Avenues - Spreading Truth: A Research Awareness E-Journal by IPRHSaurav Arora100% (2)
- Prostate Cancer Must Be Differentiated From: (BPH)Document4 paginiProstate Cancer Must Be Differentiated From: (BPH)Robert StolnicuÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Biotechnology AdvancesDocument24 paginiBiotechnology AdvancesKhaled Al GhaebÎncă nu există evaluări
- HANDOUT MTP046 Delirium PreventionDocument19 paginiHANDOUT MTP046 Delirium PreventionShyama SharmaÎncă nu există evaluări
- Therapeutic Ultrasound in Dentistry PDFDocument96 paginiTherapeutic Ultrasound in Dentistry PDFGokul SivaÎncă nu există evaluări
- Bruzzaniti2006 Articulo OsteoclastosDocument17 paginiBruzzaniti2006 Articulo OsteoclastosL Andrea CardenasÎncă nu există evaluări
- Biomedicines: Emerging Medical Treatments For Meningioma in The Molecular EraDocument21 paginiBiomedicines: Emerging Medical Treatments For Meningioma in The Molecular EraNorAmalinaÎncă nu există evaluări
- Proteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)Document306 paginiProteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)DiahaÎncă nu există evaluări
- Biodegradable Metals PDFDocument202 paginiBiodegradable Metals PDFvw5ohgjr100% (1)
- BM 1-6 MechanotransductionDocument28 paginiBM 1-6 MechanotransductionSanthoshi Sadhanaa SankarÎncă nu există evaluări
- Neuromuscular Disorders I To 12Document280 paginiNeuromuscular Disorders I To 12Mário MachadoÎncă nu există evaluări
- 8 Gingiva and Orthodontic TreatmentDocument15 pagini8 Gingiva and Orthodontic TreatmentFernanda Corrêa FumagalliÎncă nu există evaluări
- Stem Cells PicturesDocument18 paginiStem Cells Pictures奕蓁陳Încă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Metastasis MechanismsDocument16 paginiMetastasis Mechanismsivana karina peñaranda vegaÎncă nu există evaluări