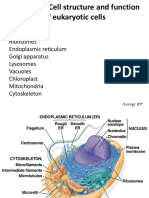
S-ar putea să vă placă și
- I Notes RETINA PDFDocument251 paginiI Notes RETINA PDFFelipe renquenÎncă nu există evaluări
- Restriction Enzymes Restriction Enzymes: Dr. P. Balaji MGR College, HosurDocument31 paginiRestriction Enzymes Restriction Enzymes: Dr. P. Balaji MGR College, HosurBalaji PaulrajÎncă nu există evaluări
- RFLP & RapdDocument25 paginiRFLP & RapdBalaji Prasanna KumarÎncă nu există evaluări
- Sandlewood Health BenefitsDocument7 paginiSandlewood Health BenefitskalaimaniÎncă nu există evaluări
- Confocal MicrosDocument33 paginiConfocal Microsfatema100% (2)
- Bio 106 Lecture 5 Chemical Basis of Heredit1Document16 paginiBio 106 Lecture 5 Chemical Basis of Heredit1Evan Jane JumamilÎncă nu există evaluări
- Chapter 6 - Cell Structure and FunctionDocument21 paginiChapter 6 - Cell Structure and FunctionSherry CarentonaÎncă nu există evaluări
- MutationDocument33 paginiMutationarslan akramÎncă nu există evaluări
- Mutations ClassificationDocument25 paginiMutations ClassificationEdessa G. FlordelizÎncă nu există evaluări
- ApoptosisDocument23 paginiApoptosisLavanya Kalapala100% (1)
- Affinity ChromatographyDocument3 paginiAffinity ChromatographyNithya Ram100% (1)
- Big Science Competition Practise Questions Years 11 - 12Document4 paginiBig Science Competition Practise Questions Years 11 - 12Happy4314Încă nu există evaluări
- Cellular RespirationDocument11 paginiCellular RespirationNiña Jennifer Potter Vergara100% (2)
- DNA Manipulative EnzymesDocument17 paginiDNA Manipulative EnzymesZain Ul AbedienÎncă nu există evaluări
- Cell Cycle Quiz-1Document3 paginiCell Cycle Quiz-1api-315443316Încă nu există evaluări
- Gene Regulation of Prokaryotic TranscriptionDocument17 paginiGene Regulation of Prokaryotic TranscriptionAyu Tri AgustinÎncă nu există evaluări
- Regulation of Gene ExpressionDocument34 paginiRegulation of Gene ExpressionJay Fickle100% (1)
- Lec1 - Biosafety and Waste Management in Molecular LabDocument47 paginiLec1 - Biosafety and Waste Management in Molecular LabClaire GonoÎncă nu există evaluări
- Protein Sorting and TransportDocument17 paginiProtein Sorting and TransportMichiko BernardoÎncă nu există evaluări
- Post-Translational Modifications To ProteinsDocument26 paginiPost-Translational Modifications To Proteinssivagurunathan.P100% (3)
- Biology Investigatory ProjectDocument15 paginiBiology Investigatory ProjectVikas Ojha69% (16)
- BIOS 302 Restriction Enzymes and DNA Modifying Enzymes PDFDocument12 paginiBIOS 302 Restriction Enzymes and DNA Modifying Enzymes PDFGauthier Djekilamber NendodoumÎncă nu există evaluări
- GE IV TH SemDocument185 paginiGE IV TH Semrahul vivekÎncă nu există evaluări
- Membrane TraffickingDocument74 paginiMembrane Traffickinganuj2907Încă nu există evaluări
- Measure of Central TendencyDocument16 paginiMeasure of Central Tendencydr.neupane27Încă nu există evaluări
- Dna Mutation & Repair MechanismDocument23 paginiDna Mutation & Repair MechanismOsama Bin RizwanÎncă nu există evaluări
- Polymerase Chain ReactionDocument22 paginiPolymerase Chain ReactionPoornaBasuri100% (1)
- Cell Adhesion MoleculesDocument14 paginiCell Adhesion MoleculesSecret Agent100% (1)
- Atomic Force MicrosDocument26 paginiAtomic Force MicrosGnana Sri Durga ParuchuriÎncă nu există evaluări
- Measuring Protein Concentration Using Absorbance at 280 NM PDFDocument3 paginiMeasuring Protein Concentration Using Absorbance at 280 NM PDFmnstnÎncă nu există evaluări
- Immunohistochemistry Principles Uses and MethodsDocument10 paginiImmunohistochemistry Principles Uses and MethodsKailash PrajapatÎncă nu există evaluări
- Sequecing - Khan AcademyDocument16 paginiSequecing - Khan AcademyMaryem SafdarÎncă nu există evaluări
- Ch-Immobilization of Enzyme PDFDocument92 paginiCh-Immobilization of Enzyme PDFVÎncă nu există evaluări
- Application of rDNA in Animal Cell Culture (Animal Biotech)Document64 paginiApplication of rDNA in Animal Cell Culture (Animal Biotech)jithinnxÎncă nu există evaluări
- Intracellular TransportDocument66 paginiIntracellular Transportalvitakhoridatul100% (1)
- Numericals On DNA TopologyDocument5 paginiNumericals On DNA TopologySadab KhanÎncă nu există evaluări
- Recombinant DNA Technology LectureDocument50 paginiRecombinant DNA Technology LectureJean Carmelette BalalloÎncă nu există evaluări
- Csi Lab Report 2Document3 paginiCsi Lab Report 2api-374131693Încă nu există evaluări
- Microscope: Micrometry and The Microbial WorldDocument3 paginiMicroscope: Micrometry and The Microbial WorldPatricia Hariramani100% (1)
- DNA Topology Supercoiling and Linking PDFDocument5 paginiDNA Topology Supercoiling and Linking PDFmanoj_rkl_07Încă nu există evaluări
- Internship Evaluation Form NewDocument3 paginiInternship Evaluation Form Newsubhan sibghatÎncă nu există evaluări
- Mechanism of Protein Targeting Into Endoplasmic ReticulumDocument24 paginiMechanism of Protein Targeting Into Endoplasmic ReticulumRebati Raman PandaÎncă nu există evaluări
- Utation: Big PictureDocument2 paginiUtation: Big PicturehomamunfatÎncă nu există evaluări
- Biology Genetic Mutation NotesDocument1 paginăBiology Genetic Mutation NotesTiffany Gallina100% (2)
- Acid RainDocument35 paginiAcid RainMajid AliÎncă nu există evaluări
- Ethnobotany NotesDocument4 paginiEthnobotany NotesBereket Yitayaw100% (1)
- Purification of DNADocument14 paginiPurification of DNAalivetutorsÎncă nu există evaluări
- DNA DamageDocument3 paginiDNA DamageAnonymous yNWMB80ffFÎncă nu există evaluări
- Staining Techniques in Biological StudiesDocument52 paginiStaining Techniques in Biological StudiesRamyaAbhishekShenoyÎncă nu există evaluări
- A Review of Methods For The Detection of Pathogenic MicroorganismsDocument16 paginiA Review of Methods For The Detection of Pathogenic MicroorganismsLuisÎncă nu există evaluări
- Basic Science of Wound HealingDocument4 paginiBasic Science of Wound HealingMarnia SulfianaÎncă nu există evaluări
- BotanyDocument18 paginiBotanyAditya RudraÎncă nu există evaluări
- Real-Time Polymerase Chain Reaction - Wikipedia, The Free EncyclopediaDocument10 paginiReal-Time Polymerase Chain Reaction - Wikipedia, The Free EncyclopediaMondal RajuÎncă nu există evaluări
- EnzymesDocument40 paginiEnzymesAmal100% (2)
- Red Biotechnology ProjectDocument5 paginiRed Biotechnology ProjectNeveen Kandil100% (2)
- Genetics and InheritanceDocument33 paginiGenetics and InheritanceLawrence Onthuga100% (1)
- Chromatin RemodelingDocument5 paginiChromatin RemodelingRohit GargÎncă nu există evaluări
- C Value ParadoxDocument4 paginiC Value ParadoxEr Purushottam PalÎncă nu există evaluări
- A Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate IndependentlyDocument5 paginiA Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate Independentlyyaqoob008Încă nu există evaluări
- Transposons in HumansDocument22 paginiTransposons in HumansRishiÎncă nu există evaluări
- Cell Structure and Functions PPT 4Document22 paginiCell Structure and Functions PPT 4rajesh duaÎncă nu există evaluări
- Extensions To MendelismDocument20 paginiExtensions To MendelismJason RodmanÎncă nu există evaluări
- Cell CycleDocument9 paginiCell Cyclestevensb055Încă nu există evaluări
- cDNA Libraries and Gene CloningDocument8 paginicDNA Libraries and Gene CloningRoberto RomeroÎncă nu există evaluări
- EG1108 RectifiersDocument14 paginiEG1108 RectifiersEunicePeñaflorÎncă nu există evaluări
- AssignmentDocument1 paginăAssignmentnaveenÎncă nu există evaluări
- Circuit Diagram: B1 (9v) Photo Transistor (L14G1 OR OP505A)Document1 paginăCircuit Diagram: B1 (9v) Photo Transistor (L14G1 OR OP505A)naveenÎncă nu există evaluări
- BIO AimDocument1 paginăBIO AimnaveenÎncă nu există evaluări
- SESSION 54c - Right Manners Positive AttitudeDocument14 paginiSESSION 54c - Right Manners Positive AttitudenaveenÎncă nu există evaluări
- Prefabricated 110303031212 Phpapp02Document119 paginiPrefabricated 110303031212 Phpapp02Sheikh Nouman Mohsin RamziÎncă nu există evaluări
- EDCLABDocument80 paginiEDCLABAmbikaÎncă nu există evaluări
- Prefab ReportDocument8 paginiPrefab ReportnaveenÎncă nu există evaluări
- 13 743 PDFDocument15 pagini13 743 PDFshardasiddhÎncă nu există evaluări
- To Study The About The Microbes Around UsDocument1 paginăTo Study The About The Microbes Around UsnaveenÎncă nu există evaluări
- Formulae Integration 1Document1 paginăFormulae Integration 1naveenÎncă nu există evaluări
- Neet Test Nov 13Document3 paginiNeet Test Nov 13naveenÎncă nu există evaluări
- Science XDocument5 paginiScience XnaveenÎncă nu există evaluări
- Bile Esculine TestDocument2 paginiBile Esculine TestVincent OngÎncă nu există evaluări
- Traditional Indian MedicineDocument3 paginiTraditional Indian MedicineShashvita RatraÎncă nu există evaluări
- Presentation List Language Disorders in ChildrenDocument3 paginiPresentation List Language Disorders in Childrenramandeep kaurÎncă nu există evaluări
- Applying Mendel's Principles: Lesson ObjectivesDocument4 paginiApplying Mendel's Principles: Lesson ObjectivesTae McLoughlinÎncă nu există evaluări
- Cervical Speculum ExaminationDocument41 paginiCervical Speculum ExaminationEdwin Delos Reyes AbuÎncă nu există evaluări
- CefuroximeDocument14 paginiCefuroximeBentoys StreetÎncă nu există evaluări
- CENE Course Outline On Psychiatric NursingDocument3 paginiCENE Course Outline On Psychiatric NursingJohn Ryan BuenaventuraÎncă nu există evaluări
- De So 3Document6 paginiDe So 3To Minh PhuongÎncă nu există evaluări
- The Nurse Leader's Role: Crisis ManagementDocument3 paginiThe Nurse Leader's Role: Crisis Managementravsab GaikwadÎncă nu există evaluări
- Winter 2010 Natural FarmerDocument48 paginiWinter 2010 Natural FarmerV1xWqÎncă nu există evaluări
- Neuromuscular Cardiovascular Hypocalcemia: Hypermagnesemia (Symptoms)Document2 paginiNeuromuscular Cardiovascular Hypocalcemia: Hypermagnesemia (Symptoms)Glormina Asprec AvenaÎncă nu există evaluări
- Labiomandibular Paresthesia Caused by Endodontic Treatment - An Anatomic and Clinical Study PDFDocument13 paginiLabiomandibular Paresthesia Caused by Endodontic Treatment - An Anatomic and Clinical Study PDFAlexandra DumitracheÎncă nu există evaluări
- Pyometra in A Cat: A Clinical Case Report: November 2021Document7 paginiPyometra in A Cat: A Clinical Case Report: November 2021Ronny Alberto GallegoÎncă nu există evaluări
- # Category Image Product Name Producer Package Hs Code Remark Key FeaturesDocument9 pagini# Category Image Product Name Producer Package Hs Code Remark Key Featuresiq_dianaÎncă nu există evaluări
- Notice: Agency Information Collection Activities Proposals, Submissions, and ApprovalsDocument1 paginăNotice: Agency Information Collection Activities Proposals, Submissions, and ApprovalsJustia.comÎncă nu există evaluări
- Test Name Result Unit Reference Interval EndocrinolgyDocument1 paginăTest Name Result Unit Reference Interval Endocrinolgygeetha kodaliÎncă nu există evaluări
- 536 BPG Assessment Foot Ulcer 1Document115 pagini536 BPG Assessment Foot Ulcer 1FebricaPutriArthaÎncă nu există evaluări
- Presentation 1Document12 paginiPresentation 1Rizka FarahinÎncă nu există evaluări
- Phytochemical Analysis, Antimicrobial and Antioxidant Activities of Euphorbia Juss.) : An Unexplored Medicinal Herb Reported From CameroonDocument15 paginiPhytochemical Analysis, Antimicrobial and Antioxidant Activities of Euphorbia Juss.) : An Unexplored Medicinal Herb Reported From CameroonClara JamirÎncă nu există evaluări
- Efficacy of Doctorvox On Mutational FalsettoDocument8 paginiEfficacy of Doctorvox On Mutational FalsettoANA CRISTINA MENDEZ DIAZÎncă nu există evaluări
- Areport 08Document247 paginiAreport 08Mithilesh JhaÎncă nu există evaluări
- Employers Guide - InteractiveDocument24 paginiEmployers Guide - Interactiveapi-285398586Încă nu există evaluări
- A Epithelial - Tissue1 16 12 14Document32 paginiA Epithelial - Tissue1 16 12 14Abdulaziz AbdullahiÎncă nu există evaluări
- Cardiovascular Responses To An Isometric Handgrip Exercise in Females With PrehypertensionDocument7 paginiCardiovascular Responses To An Isometric Handgrip Exercise in Females With PrehypertensionGuilherme LimaÎncă nu există evaluări
- Abreviaciones ChatDocument4 paginiAbreviaciones ChatrojarodÎncă nu există evaluări
- Dairy SpecialDocument24 paginiDairy Specialthanh ba matÎncă nu există evaluări
- ABC of Sexual Health: Taking A Sexual HistoryDocument10 paginiABC of Sexual Health: Taking A Sexual HistoryCristina PerezÎncă nu există evaluări