S-ar putea să vă placă și
- Hologramas Luz y Armonia PeruDocument8 paginiHologramas Luz y Armonia PeruEl Olivo InmueblesÎncă nu există evaluări
- Dientes HistologíaDocument12 paginiDientes HistologíaJulio Loján100% (1)
- Tiburones Rayas y Quimeras en MexicoDocument6 paginiTiburones Rayas y Quimeras en MexicoLaísa SalanderÎncă nu există evaluări
- Frases y Palabras LlanerasDocument25 paginiFrases y Palabras Llanerasjuliandcj100% (1)
- La CochinillaDocument6 paginiLa Cochinillatou cheÎncă nu există evaluări
- Biodiv 108 Art 1Document6 paginiBiodiv 108 Art 1Diana BarrónÎncă nu există evaluări
- Biodiv 108 Art 2Document5 paginiBiodiv 108 Art 2Diana BarrónÎncă nu există evaluări
- Biodiv110art4 PDFDocument3 paginiBiodiv110art4 PDFDiana BarrónÎncă nu există evaluări
- Biodiv 111 Art 3Document5 paginiBiodiv 111 Art 3Diana BarrónÎncă nu există evaluări
- Biodiv109art2 PDFDocument5 paginiBiodiv109art2 PDFDiana BarrónÎncă nu există evaluări
- Biodiv112art3 PDFDocument7 paginiBiodiv112art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv109art1 PDFDocument6 paginiBiodiv109art1 PDFDiana Barrón100% (1)
- Biodiv108art3 PDFDocument5 paginiBiodiv108art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv109art3b PDFDocument5 paginiBiodiv109art3b PDFDiana BarrónÎncă nu există evaluări
- Biodiv 110 Art 2Document4 paginiBiodiv 110 Art 2Diana BarrónÎncă nu există evaluări
- Biodiv 110 Art 3Document4 paginiBiodiv 110 Art 3Diana BarrónÎncă nu există evaluări
- Biodiv112art2 PDFDocument4 paginiBiodiv112art2 PDFDiana BarrónÎncă nu există evaluări
- Biodiv 112 Art 1Document5 paginiBiodiv 112 Art 1Diana BarrónÎncă nu există evaluări
- Biodiv113art3 PDFDocument5 paginiBiodiv113art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv 113 Art 2Document4 paginiBiodiv 113 Art 2Diana BarrónÎncă nu există evaluări
- Biodiv 114 Art 2Document5 paginiBiodiv 114 Art 2Diana BarrónÎncă nu există evaluări
- Lectura Biodiversidad PDFDocument7 paginiLectura Biodiversidad PDFMayer ArizaÎncă nu există evaluări
- Biodiv114art3 PDFDocument4 paginiBiodiv114art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv 114 Art 1Document7 paginiBiodiv 114 Art 1Diana BarrónÎncă nu există evaluări
- Biodiv115art1 PDFDocument6 paginiBiodiv115art1 PDFDiana BarrónÎncă nu există evaluări
- Biodiv115art1 PDFDocument6 paginiBiodiv115art1 PDFDiana BarrónÎncă nu există evaluări
- Biodiv116art3 PDFDocument5 paginiBiodiv116art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv115art2 PDFDocument5 paginiBiodiv115art2 PDFDiana BarrónÎncă nu există evaluări
- Biodiv115art2 PDFDocument5 paginiBiodiv115art2 PDFDiana BarrónÎncă nu există evaluări
- Biodiv119art2 PDFDocument5 paginiBiodiv119art2 PDFDiana BarrónÎncă nu există evaluări
- Biodiv115art3 PDFDocument5 paginiBiodiv115art3 PDFDiana BarrónÎncă nu există evaluări
- Biodiv116art1 PDFDocument6 paginiBiodiv116art1 PDFDiana BarrónÎncă nu există evaluări
- Biodiv 116 Art 2Document5 paginiBiodiv 116 Art 2Diana BarrónÎncă nu există evaluări
- Biodiv117ar1 PDFDocument6 paginiBiodiv117ar1 PDFDiana BarrónÎncă nu există evaluări
- Enlace QuímicoDocument6 paginiEnlace QuímicoWilmer John Quincho CabreraÎncă nu există evaluări
- Cuestionario de EgoDocument15 paginiCuestionario de EgoMel alixon Farell AguilarÎncă nu există evaluări
- Fertilizantes Plaguicidas Vigilados Mar 2013 Productores ImpoDocument198 paginiFertilizantes Plaguicidas Vigilados Mar 2013 Productores ImpoLaura CelisÎncă nu există evaluări
- Practica Transferencia de Oxígeno Bioingeniería 2Document17 paginiPractica Transferencia de Oxígeno Bioingeniería 2EduardoÎncă nu există evaluări
- Hematopoyesis FetalDocument20 paginiHematopoyesis Fetalmanduca420Încă nu există evaluări
- Winalite Mexico PresentacionDocument54 paginiWinalite Mexico PresentaciontetewinaliteÎncă nu există evaluări
- 72 Sanmiguel - Callejon SucreDocument63 pagini72 Sanmiguel - Callejon SucreJuariDocs100% (3)
- Informe Bioquimica PapaDocument10 paginiInforme Bioquimica PapaGerardo Mogollon RamosÎncă nu există evaluări
- 01 BoliviaDocument84 pagini01 BoliviaPaolita Pinzón ArangoÎncă nu există evaluări
- Variedades de PapaDocument1 paginăVariedades de PapaJuan lo llamaban y se apodaba CharrasqueadoÎncă nu există evaluări
- Repaso Biología MolecularDocument10 paginiRepaso Biología MolecularRodrigo RodríguezÎncă nu există evaluări
- Biologia j2015Document11 paginiBiologia j2015carla soaresÎncă nu există evaluări
- 2.1 Biomoleculas y BioelemntosDocument19 pagini2.1 Biomoleculas y BioelemntoskarlaÎncă nu există evaluări
- 8 Asignatura III 8 T R A L U C eDocument25 pagini8 Asignatura III 8 T R A L U C eEzequiel Luppi0% (1)
- AntiviralesDocument44 paginiAntiviralesIngrid Romina Rojas ChamorroÎncă nu există evaluări
- Peec 2011-Hematologia PDFDocument21 paginiPeec 2011-Hematologia PDFErick2007Încă nu există evaluări
- Caso Clínico de Intregración Hemodinámica y Electrica 2Document17 paginiCaso Clínico de Intregración Hemodinámica y Electrica 2Natalia RodríguezÎncă nu există evaluări
- Lectura Complementaria 2Document4 paginiLectura Complementaria 2c baÎncă nu există evaluări
- Apuntes FisiologiaDocument29 paginiApuntes Fisiologiamgilermo1Încă nu există evaluări
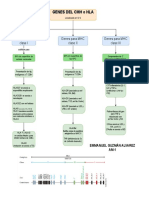
- Genes de HLA o MHCDocument1 paginăGenes de HLA o MHCEmmanuel GÎncă nu există evaluări
- El Procesado de Frutas y VerdurasDocument7 paginiEl Procesado de Frutas y VerdurasLos Viajes De Tolomeo TeatroÎncă nu există evaluări
- Cripreservación Bagre Rayado PDFDocument113 paginiCripreservación Bagre Rayado PDFLizbeth Zuta PinedoÎncă nu există evaluări
- Mono 11 Ecorregiones CDocument10 paginiMono 11 Ecorregiones CJorge RosalesÎncă nu există evaluări
- Informe Bioquímica Respiración TisularDocument7 paginiInforme Bioquímica Respiración TisularEduardo Ramírez CapristánÎncă nu există evaluări
- Electricidad Automotriz BasicaDocument329 paginiElectricidad Automotriz BasicaFranco Jamil Alarcón ApazaÎncă nu există evaluări
- Aap C05 U1 EvPro 1a5Document2 paginiAap C05 U1 EvPro 1a5Lizeth CubillosÎncă nu există evaluări