S-ar putea să vă placă și
- Recent Developments in the Chemistry of Natural Phenolic Compounds: Proceedings of the Plant Phenolics Group SymposiumDe la EverandRecent Developments in the Chemistry of Natural Phenolic Compounds: Proceedings of the Plant Phenolics Group SymposiumW. D. OllisÎncă nu există evaluări
- Properties of StarchDocument13 paginiProperties of StarchPadmavati KarthicÎncă nu există evaluări
- References For StarchDocument2 paginiReferences For Starchapi-388978795Încă nu există evaluări
- Supercritical Fluid Cleaning: Fundamentals, Technology and ApplicationsDe la EverandSupercritical Fluid Cleaning: Fundamentals, Technology and ApplicationsSamuel P. SawanÎncă nu există evaluări
- Gelatinization Mechanism of STARCH GRANULESDocument6 paginiGelatinization Mechanism of STARCH GRANULESVic KeyÎncă nu există evaluări
- A Treatise on Adulterations of Food, and Culinary Poisons Exhibiting the Fraudulent Sophistications of Bread, Beer, Wine, Spiritous Liquors, Tea, Coffee, Cream, Confectionery, Vinegar, Mustard, Pepper, Cheese, Olive Oil, Pickles, and Other Articles Employed in Domestic EconomyDe la EverandA Treatise on Adulterations of Food, and Culinary Poisons Exhibiting the Fraudulent Sophistications of Bread, Beer, Wine, Spiritous Liquors, Tea, Coffee, Cream, Confectionery, Vinegar, Mustard, Pepper, Cheese, Olive Oil, Pickles, and Other Articles Employed in Domestic EconomyÎncă nu există evaluări
- Native Wheat Starch Info SheetDocument4 paginiNative Wheat Starch Info SheetBuia DanutÎncă nu există evaluări
- Starch GelatinizationDocument1 paginăStarch GelatinizationFaleh Setia BudiÎncă nu există evaluări
- Chemical Changes During Extrusion Cooking - Camire PDFDocument13 paginiChemical Changes During Extrusion Cooking - Camire PDFmarmaduke32Încă nu există evaluări
- Cow Urine Guide Provides Health BenefitsDocument34 paginiCow Urine Guide Provides Health BenefitsSurendraBaba50% (2)
- Plant LectinsDocument25 paginiPlant Lectinsshady182100% (1)
- A Fresh Take On Fake MeatDocument3 paginiA Fresh Take On Fake MeatMark Francis PlazaÎncă nu există evaluări
- Is 3965 PDFDocument13 paginiIs 3965 PDFYuvaraj SathishÎncă nu există evaluări
- Starch. Advances in Structure and Function - T. L. Barsby PDFDocument233 paginiStarch. Advances in Structure and Function - T. L. Barsby PDFGrigore VladÎncă nu există evaluări
- Axially Splitcase Pumps - HS & HST SeriesDocument12 paginiAxially Splitcase Pumps - HS & HST SeriesSuhas Natu100% (1)
- Starch GranuleDocument9 paginiStarch GranuleKay Buchorchor JnrÎncă nu există evaluări
- Itenary Trip To Thailand 2018 (Fix)Document13 paginiItenary Trip To Thailand 2018 (Fix)Riza ArindaÎncă nu există evaluări
- InTech-Nutritional Value of Soybean MealDocument21 paginiInTech-Nutritional Value of Soybean MealGuillermo UCÎncă nu există evaluări
- Carbohydrates Structure and NomenclatureDocument60 paginiCarbohydrates Structure and NomenclatureNikka Mia AbadiesÎncă nu există evaluări
- BIO20 - The Biomolecules (Handouts) PDFDocument13 paginiBIO20 - The Biomolecules (Handouts) PDFKeith Ryan LapizarÎncă nu există evaluări
- 2B Carbs and LipidsDocument26 pagini2B Carbs and LipidsShawn NagraÎncă nu există evaluări
- The Structure and Function of MacromoleculesDocument70 paginiThe Structure and Function of MacromoleculesAhmadMaulanaIfanAkbasÎncă nu există evaluări
- Ref G2 PolysaccharideDocument41 paginiRef G2 PolysaccharideLEE ZHI XUAN A21SC0118Încă nu există evaluări
- Introductory Biochemistry: CarbohydratesDocument68 paginiIntroductory Biochemistry: CarbohydratesMahrukh SaeedÎncă nu există evaluări
- Dr.H.Mohammad Hanafi, Mbbs (Syd.) .MS.: Fakultas Kedokteran UnairDocument48 paginiDr.H.Mohammad Hanafi, Mbbs (Syd.) .MS.: Fakultas Kedokteran UnairahmustofaÎncă nu există evaluări
- Lec - 1 - CarbohydrateDocument19 paginiLec - 1 - CarbohydrateFjÎncă nu există evaluări
- NCERT Solutions For Class 12 Chemistry Chapter 14 BiomoleculesDocument7 paginiNCERT Solutions For Class 12 Chemistry Chapter 14 BiomoleculesjeyavegneshÎncă nu există evaluări
- Ch05 MacromoleculesCampbellDocument69 paginiCh05 MacromoleculesCampbellCarl Jasper AquinoÎncă nu există evaluări
- Biomolecules: Agitha R. Menon PGT AECS, IndoreDocument19 paginiBiomolecules: Agitha R. Menon PGT AECS, Indoreshivanigangwar69Încă nu există evaluări
- Carbohydrates 1: MSL: BiochemistryDocument50 paginiCarbohydrates 1: MSL: Biochemistryنجوي عبدالوهابÎncă nu există evaluări
- Carbohydrates: Molecular BiochemistryDocument36 paginiCarbohydrates: Molecular BiochemistryDozdiÎncă nu există evaluări
- Biokimia1 CarbohydratesDocument52 paginiBiokimia1 CarbohydratesfinaÎncă nu există evaluări
- 1 ChoDocument79 pagini1 ChoFrance Jan First SaplacoÎncă nu există evaluări
- Monosacharides 2015Document94 paginiMonosacharides 2015Syeda AlizaÎncă nu există evaluări
- Carbohydrates ( - Cho / - C O)Document55 paginiCarbohydrates ( - Cho / - C O)nemidoonamÎncă nu există evaluări
- Carbohydrates (Monosaccharides) : Anbar University-College of Pharmacy-Clinical Laboratory Sciences Department 2019-2020Document22 paginiCarbohydrates (Monosaccharides) : Anbar University-College of Pharmacy-Clinical Laboratory Sciences Department 2019-2020Zahid Yaqoob BhatÎncă nu există evaluări
- Carbohidratos 4: Jorge A Durán VDocument22 paginiCarbohidratos 4: Jorge A Durán VSebastián ArenasÎncă nu există evaluări
- Carbs: Sources of Energy, Structures & FunctionsDocument28 paginiCarbs: Sources of Energy, Structures & FunctionsKiruthikasweetyÎncă nu există evaluări
- 202003291612341624kuaum Yadav Structure and Properties of CarbohydratesDocument8 pagini202003291612341624kuaum Yadav Structure and Properties of CarbohydratesyasmienmiguelÎncă nu există evaluări
- Chapter 5 The Structue and Function of MacromoleculesDocument71 paginiChapter 5 The Structue and Function of MacromoleculesKhoirunnisa LuthfiyyahÎncă nu există evaluări
- Basic Biology: Carbohydrates and Cell OrganisationDocument26 paginiBasic Biology: Carbohydrates and Cell OrganisationZoé LennonÎncă nu există evaluări
- Classification of CarbohydratesDocument97 paginiClassification of CarbohydratesALEM, RALPH LAURENCE B.Încă nu există evaluări
- Wa0013.Document30 paginiWa0013.VekeMalaÎncă nu există evaluări
- 9 CarbohydratesDocument37 pagini9 Carbohydratesjaya1129Încă nu există evaluări
- Chapter 3 Biological MacromoleculesDocument32 paginiChapter 3 Biological MacromoleculeslolaÎncă nu există evaluări
- The Nature of Carbohydrates PDFDocument65 paginiThe Nature of Carbohydrates PDFShahriarÎncă nu există evaluări
- Lecture 2 Cell BiologyDocument27 paginiLecture 2 Cell Biologybobo-girl123Încă nu există evaluări
- Carbohydrates ASAS 4104Document53 paginiCarbohydrates ASAS 4104AlbanMugotiÎncă nu există evaluări
- Karbohidrat JupeDocument47 paginiKarbohidrat JupeRayanda AdiputraÎncă nu există evaluări
- Carob Hydrates SummaryDocument17 paginiCarob Hydrates Summaryalizay91Încă nu există evaluări
- Carbohydrates: Energy Sources and Building BlocksDocument34 paginiCarbohydrates: Energy Sources and Building BlocksUrooj GulÎncă nu există evaluări
- Carbohydrates: Structure and FunctionsDocument47 paginiCarbohydrates: Structure and FunctionsNathanael UlrichsternÎncă nu există evaluări
- BiomoleculesDocument32 paginiBiomoleculesanumeha.kinsui23Încă nu există evaluări
- Chem Board FINAL (00) EditedDocument23 paginiChem Board FINAL (00) Editedattishya1104Încă nu există evaluări
- Carbohydrate: ClassificationDocument67 paginiCarbohydrate: ClassificationKim Ryan Ello CagasÎncă nu există evaluări
- Lipids PDFDocument6 paginiLipids PDFGulam Muhammad AkhtarÎncă nu există evaluări
- UNIT-14 BIOMOLECULES 2023Document22 paginiUNIT-14 BIOMOLECULES 2023jagannathanÎncă nu există evaluări
- FC Lect Topic4d AY17-18 StudentDocument29 paginiFC Lect Topic4d AY17-18 StudentRaysonChooÎncă nu există evaluări
- CarbsDocument34 paginiCarbsManendra PatwaÎncă nu există evaluări
- PRC Authenication Form PDFDocument2 paginiPRC Authenication Form PDFMichael Densing InsoÎncă nu există evaluări
- 10 1067@j Cpradiol 2009 07 005 PDFDocument19 pagini10 1067@j Cpradiol 2009 07 005 PDFJess LejardeÎncă nu există evaluări
- Thrombosis Research: Review ArticleDocument8 paginiThrombosis Research: Review ArticleJess LejardeÎncă nu există evaluări
- Abdominal AneurysmDocument3 paginiAbdominal AneurysmJess LejardeÎncă nu există evaluări
- Thrombosis Research: Review ArticleDocument8 paginiThrombosis Research: Review ArticleJess LejardeÎncă nu există evaluări
- KAP Study Guideline for Community OphthalmologyDocument3 paginiKAP Study Guideline for Community OphthalmologySidik Syah PutraÎncă nu există evaluări
- Philippine Guidelines on Diagnosis and Treatment of Community-Acquired PneumoniaDocument13 paginiPhilippine Guidelines on Diagnosis and Treatment of Community-Acquired Pneumoniamits98Încă nu există evaluări
- AD Antiseptics PDFDocument11 paginiAD Antiseptics PDFJess LejardeÎncă nu există evaluări
- Lymphatic System ReviewerDocument49 paginiLymphatic System ReviewerJess Lejarde100% (1)
- Exp 2 Protein DeterminationDocument5 paginiExp 2 Protein DeterminationNur Fadhilah100% (1)
- 1229.1 Steam Sterilization by Direct ContactDocument3 pagini1229.1 Steam Sterilization by Direct ContactWasimÎncă nu există evaluări
- AJODO Eliades2006 - Orthodontic Materials Research and Applications Part 1-Current Status and Projected Future Developments in Bonding and Adhesives.Document7 paginiAJODO Eliades2006 - Orthodontic Materials Research and Applications Part 1-Current Status and Projected Future Developments in Bonding and Adhesives.drgeorgejose7818Încă nu există evaluări
- Bref Polymer EsDocument319 paginiBref Polymer EsMaFher JimenezÎncă nu există evaluări
- Aws Gtaw Study GuideDocument2 paginiAws Gtaw Study GuideTouil HoussemÎncă nu există evaluări
- Math 6 Unit 8 Volume of Solids and LiquidsDocument1 paginăMath 6 Unit 8 Volume of Solids and LiquidsRobi AkmalÎncă nu există evaluări
- Quantum-Mechanical Aspects of The L. Pauling's Resonance Theory.Document4 paginiQuantum-Mechanical Aspects of The L. Pauling's Resonance Theory.Bezverkhniy VolodymyrÎncă nu există evaluări
- Colder Products Company Full CatalogDocument166 paginiColder Products Company Full CatalogDENNIZÎncă nu există evaluări
- Recycling of Pad-Batch Washing Textile Wastewater Through Advanced Oxidation Processes and Its Reusability Assessment For Turkish Textile IndustDocument7 paginiRecycling of Pad-Batch Washing Textile Wastewater Through Advanced Oxidation Processes and Its Reusability Assessment For Turkish Textile IndustGizem D.Încă nu există evaluări
- 11 Numerical AnalysisDocument9 pagini11 Numerical Analysisعزالدين حسنÎncă nu există evaluări
- Boiler Efficiency Calculations SheetDocument5 paginiBoiler Efficiency Calculations Sheetkalyanm20351578% (9)
- SodaPDF-converted-Exercise No. 7 - Soil Sample Collection and PreparationDocument30 paginiSodaPDF-converted-Exercise No. 7 - Soil Sample Collection and PreparationJacky Lou GermanoÎncă nu există evaluări
- PESSAT-Chemistry-Model-PaperDocument12 paginiPESSAT-Chemistry-Model-PaperpullagalkÎncă nu există evaluări
- On Water Cycle With Unlocking of DifficultiesDocument38 paginiOn Water Cycle With Unlocking of DifficultiesShirly Basilio100% (1)
- Problem #2b: Chromium Crystallizes With A Body-Centered Cubic Unit Cell. The Radius of ADocument8 paginiProblem #2b: Chromium Crystallizes With A Body-Centered Cubic Unit Cell. The Radius of ARadica AyuÎncă nu există evaluări
- An Updated Review On IPQC Tests For Sterile and Non Sterile ProductsDocument12 paginiAn Updated Review On IPQC Tests For Sterile and Non Sterile ProductsJvnÎncă nu există evaluări
- What's New - PV Elite 2018Document28 paginiWhat's New - PV Elite 2018SathiyaseelanÎncă nu există evaluări
- Full Download Engineering Applications in Sustainable Design and Development 1st Edition Striebig Solutions ManualDocument12 paginiFull Download Engineering Applications in Sustainable Design and Development 1st Edition Striebig Solutions Manualjherallapizm100% (38)
- Abaco Moody PDFDocument1 paginăAbaco Moody PDFAxo Pijo CopónÎncă nu există evaluări
- The Periodic Table of ElementsDocument39 paginiThe Periodic Table of ElementsSamKris Guerrero Malasaga100% (1)
- Booklet 2 Topic 1Document61 paginiBooklet 2 Topic 1Sarah Al ObaidanÎncă nu există evaluări
- Photosynthesis, Respiration, and Long-Distance TransportDocument2 paginiPhotosynthesis, Respiration, and Long-Distance TransportAlv1en HidayatÎncă nu există evaluări
- Documents Resources MacDermid Erifon HD856 Issue 10 A4Document2 paginiDocuments Resources MacDermid Erifon HD856 Issue 10 A4Pedro DutraÎncă nu există evaluări
- Astm A999a999mDocument12 paginiAstm A999a999mHammadÎncă nu există evaluări
- Practice 4ADocument22 paginiPractice 4ACamha NguyenÎncă nu există evaluări
- The Chemistry of Heterocycles Structure, Reactions, Syntheses, and ApplicationsDocument571 paginiThe Chemistry of Heterocycles Structure, Reactions, Syntheses, and Applications17.Hồ Nguyên Khang100% (1)
- Chap 7 PDFDocument45 paginiChap 7 PDFSuharto Masacal AmpasoÎncă nu există evaluări
- Comparison of Architectural Coating SpecificationsDocument5 paginiComparison of Architectural Coating SpecificationsvopyrupyrÎncă nu există evaluări
- Acid Cleaning For Nickel Alloys Pages From Handbook of Engineering Practice of Materials and Corrosion by Jung-Chul (Thomas) EunDocument9 paginiAcid Cleaning For Nickel Alloys Pages From Handbook of Engineering Practice of Materials and Corrosion by Jung-Chul (Thomas) EunMohammad TaherÎncă nu există evaluări
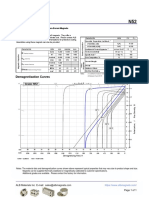
- N52 Grade Neodymium Magnets DataDocument1 paginăN52 Grade Neodymium Magnets DataSteve HsuÎncă nu există evaluări
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeDe la EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeEvaluare: 4.5 din 5 stele4.5/5 (3)
- Stuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldDe la EverandStuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldEvaluare: 4 din 5 stele4/5 (289)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeDe la EverandChemistry for Breakfast: The Amazing Science of Everyday LifeEvaluare: 4.5 din 5 stele4.5/5 (14)
- Guidelines for Asset Integrity ManagementDe la EverandGuidelines for Asset Integrity ManagementEvaluare: 5 din 5 stele5/5 (1)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeDe la EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeEvaluare: 4 din 5 stele4/5 (9)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsDe la EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsEvaluare: 4 din 5 stele4/5 (146)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeDe la EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeEvaluare: 5 din 5 stele5/5 (1)
- Coating and Drying Defects: Troubleshooting Operating ProblemsDe la EverandCoating and Drying Defects: Troubleshooting Operating ProblemsEvaluare: 5 din 5 stele5/5 (1)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolDe la EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolÎncă nu există evaluări
- An Introduction to the Periodic Table of Elements : Chemistry Textbook Grade 8 | Children's Chemistry BooksDe la EverandAn Introduction to the Periodic Table of Elements : Chemistry Textbook Grade 8 | Children's Chemistry BooksEvaluare: 5 din 5 stele5/5 (1)
- Guidelines for Defining Process Safety Competency RequirementsDe la EverandGuidelines for Defining Process Safety Competency RequirementsEvaluare: 3 din 5 stele3/5 (1)
- Napoleon's Buttons: 17 Molecules That Changed HistoryDe la EverandNapoleon's Buttons: 17 Molecules That Changed HistoryEvaluare: 4 din 5 stele4/5 (25)
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsDe la EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsEvaluare: 5 din 5 stele5/5 (3)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilDe la EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilEvaluare: 5 din 5 stele5/5 (1)
- It's Elemental: The Hidden Chemistry in EverythingDe la EverandIt's Elemental: The Hidden Chemistry in EverythingEvaluare: 4 din 5 stele4/5 (10)
- Meltdown: Nuclear disaster and the human cost of going criticalDe la EverandMeltdown: Nuclear disaster and the human cost of going criticalEvaluare: 5 din 5 stele5/5 (5)
- Chemical Elements Pocket Guide: Detailed Summary of the Periodic TableDe la EverandChemical Elements Pocket Guide: Detailed Summary of the Periodic TableÎncă nu există evaluări
- Chemistry: a QuickStudy Laminated Reference GuideDe la EverandChemistry: a QuickStudy Laminated Reference GuideEvaluare: 5 din 5 stele5/5 (1)
- The Periodic Table: A Very Short IntroductionDe la EverandThe Periodic Table: A Very Short IntroductionEvaluare: 4.5 din 5 stele4.5/5 (3)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeDe la EverandChemistry for Breakfast: The Amazing Science of Everyday LifeEvaluare: 4.5 din 5 stele4.5/5 (90)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableDe la EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableEvaluare: 3.5 din 5 stele3.5/5 (22)
- Introduction to Strategies for Organic SynthesisDe la EverandIntroduction to Strategies for Organic SynthesisÎncă nu există evaluări
- Guidelines for Integrating Process Safety into Engineering ProjectsDe la EverandGuidelines for Integrating Process Safety into Engineering ProjectsÎncă nu există evaluări
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeDe la EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeEvaluare: 4 din 5 stele4/5 (1)