S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Remineral - Per Ma Culture ArticleDocument4 paginiRemineral - Per Ma Culture Articlelauritsen2Încă nu există evaluări
- A Guide To Community Food ProjectsDocument19 paginiA Guide To Community Food ProjectsGulf Restoration NetworkÎncă nu există evaluări
- Bio-Char Sequestration in Terrestrial EcosystemsDocument25 paginiBio-Char Sequestration in Terrestrial EcosystemsBethAnnHunterÎncă nu există evaluări
- Converting Used Cooking Oil Into Diesel FuelDocument7 paginiConverting Used Cooking Oil Into Diesel FuelFriends of Baton Rouge Audubon SocietyÎncă nu există evaluări
- Population Growth, Warming, and Water SupplyDocument4 paginiPopulation Growth, Warming, and Water SupplyGulf Restoration NetworkÎncă nu există evaluări
- Food Recovery Resource Guide: BrochureDocument55 paginiFood Recovery Resource Guide: BrochureGreater Charlotte Harbor Sierra ClubÎncă nu există evaluări
- Remineral - Per Ma Culture ArticleDocument4 paginiRemineral - Per Ma Culture Articlelauritsen2Încă nu există evaluări
- Passive Solar 202Document32 paginiPassive Solar 202Gulf Restoration NetworkÎncă nu există evaluări
- Passive Solar ManualDocument4 paginiPassive Solar ManualGulf Restoration NetworkÎncă nu există evaluări
- How To Organize An Urban Fruit HarvestDocument26 paginiHow To Organize An Urban Fruit HarvestGulf Restoration NetworkÎncă nu există evaluări
- New Gleaning Project Prevents WasteDocument6 paginiNew Gleaning Project Prevents WasteGulf Restoration NetworkÎncă nu există evaluări
- The Use of Harvested Rainwater and GreywaterDocument1 paginăThe Use of Harvested Rainwater and GreywaterGulf Restoration NetworkÎncă nu există evaluări
- Understanding Gleaning Historical and Cultural ContextsDocument28 paginiUnderstanding Gleaning Historical and Cultural ContextsGulf Restoration NetworkÎncă nu există evaluări
- How To Start A Gleaning ProgramDocument6 paginiHow To Start A Gleaning ProgramGulf Restoration NetworkÎncă nu există evaluări
- Gleaning Crops FactsheetDocument2 paginiGleaning Crops FactsheetGulf Restoration NetworkÎncă nu există evaluări
- Adopt-a-Farm: Gleaning Project For FarmersDocument1 paginăAdopt-a-Farm: Gleaning Project For FarmersGulf Restoration NetworkÎncă nu există evaluări
- Energy Conservation For Greenhouse GrowersDocument3 paginiEnergy Conservation For Greenhouse GrowersDavidHarelÎncă nu există evaluări
- Adopt-a-Farm: Gleaning Project For FarmersDocument1 paginăAdopt-a-Farm: Gleaning Project For FarmersGulf Restoration NetworkÎncă nu există evaluări
- Green Churches of IndiaDocument15 paginiGreen Churches of IndiaGulf Restoration NetworkÎncă nu există evaluări
- Day Lighting Schools: Good Grades and Good BehavoirDocument6 paginiDay Lighting Schools: Good Grades and Good BehavoirGulf Restoration NetworkÎncă nu există evaluări
- Greywater System Design CoordinationDocument2 paginiGreywater System Design CoordinationGulf Restoration NetworkÎncă nu există evaluări
- Guide To Using Gray WaterDocument2 paginiGuide To Using Gray WaterGulf Restoration NetworkÎncă nu există evaluări
- Bio-Char Sequestration in Terrestrial EcosystemsDocument25 paginiBio-Char Sequestration in Terrestrial EcosystemsBethAnnHunterÎncă nu există evaluări
- Take Care of Oregon Days Project Ideas - 150 Project Ideas CombinedDocument150 paginiTake Care of Oregon Days Project Ideas - 150 Project Ideas CombinedSkourtosMixalisÎncă nu există evaluări
- Classroom Lighting KnowhowDocument8 paginiClassroom Lighting KnowhowPapatachiarchisEfthy100% (1)
- Ogawa: Parabolic Solar CookersDocument2 paginiOgawa: Parabolic Solar CookersGulf Restoration NetworkÎncă nu există evaluări
- Thermal Curtains - Making Your Windows WarmDocument3 paginiThermal Curtains - Making Your Windows WarmTsamparliAnastasiaÎncă nu există evaluări
- Appropriate Technology and KidsDocument2 paginiAppropriate Technology and KidsVogiatziTaniaÎncă nu există evaluări
- Parabolic Solar CookersDocument5 paginiParabolic Solar CookersTsaousisGiannisÎncă nu există evaluări
- Watt Watchers Student Energy Patrol Program ManualDocument84 paginiWatt Watchers Student Energy Patrol Program ManualGulf Restoration NetworkÎncă nu există evaluări
- Quantitative Analysis of The Relative Effectiveness of 3 Iliotibial Band StretchesDocument4 paginiQuantitative Analysis of The Relative Effectiveness of 3 Iliotibial Band Stretchessalthill01Încă nu există evaluări
- MukosaDocument3 paginiMukosaMarf PennyÎncă nu există evaluări
- Evolutiontestreviewdoc 2015Document5 paginiEvolutiontestreviewdoc 2015api-265621022Încă nu există evaluări
- Rule1 Stop TalkingDocument180 paginiRule1 Stop TalkingElKooder100% (1)
- Soal Pat Inggris Kelas 7 - 2021Document9 paginiSoal Pat Inggris Kelas 7 - 2021Bella Septiani FaryanÎncă nu există evaluări
- Presentation On Hydration PDFDocument17 paginiPresentation On Hydration PDFManuel CastroÎncă nu există evaluări
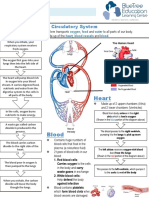
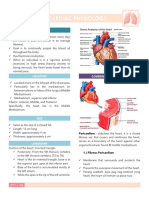
- Circulatory System Notes SLDocument1 paginăCirculatory System Notes SLRathna PalaniappanÎncă nu există evaluări
- Budget of Work Science Grade 3Document1 paginăBudget of Work Science Grade 3Mary Chovie BacusÎncă nu există evaluări
- Heart beats 100,000 times dailyDocument7 paginiHeart beats 100,000 times dailyKyle MañiboÎncă nu există evaluări
- Carbohydrates (Monosaccharides)Document56 paginiCarbohydrates (Monosaccharides)khadija100% (2)
- Its Only A PenisDocument29 paginiIts Only A PenisAdam BlairÎncă nu există evaluări
- Mzbca N2 LK 7Document342 paginiMzbca N2 LK 7Jesús Enrique Turín VilcaÎncă nu există evaluări
- Lca Mangatarem1 PDFDocument58 paginiLca Mangatarem1 PDFBea 'Jane0% (1)
- Biological Classification Class 11 Notes BiologyDocument6 paginiBiological Classification Class 11 Notes BiologyVyjayanthiÎncă nu există evaluări
- PD22 Hap1 L03Document33 paginiPD22 Hap1 L03Ka Yan LAUÎncă nu există evaluări
- Erl mf2 PDFDocument2 paginiErl mf2 PDFJorgeÎncă nu există evaluări
- The Digestive System Challenges and OpportunitiesDocument9 paginiThe Digestive System Challenges and OpportunitiesPhelipe PessanhaÎncă nu există evaluări
- Lecture 1 TeaDocument32 paginiLecture 1 Teayjonathan002Încă nu există evaluări
- Venture CityDocument34 paginiVenture CityDaniel Hidalgo VerzobiasÎncă nu există evaluări
- Solutions - AIATS Medical-2020 (XI Studying) - Test-5 (Code-C & D) - (27!01!2019)Document28 paginiSolutions - AIATS Medical-2020 (XI Studying) - Test-5 (Code-C & D) - (27!01!2019)Ûdây RäjpütÎncă nu există evaluări
- The Oak Buprestid Beetle, Agrilus Biguttatus (F.) (Col., Buprestidae), CL Recent FactorDocument5 paginiThe Oak Buprestid Beetle, Agrilus Biguttatus (F.) (Col., Buprestidae), CL Recent FactorOrgován EditÎncă nu există evaluări
- To Study The Quantity Case in Present in Different Sample of MilkDocument7 paginiTo Study The Quantity Case in Present in Different Sample of Milkbelly4u100% (2)
- Physiology - Last Minute Revision Points: DR - SreetejaDocument25 paginiPhysiology - Last Minute Revision Points: DR - SreetejaBanu KubendiranÎncă nu există evaluări
- Dialog 1: Siang, Anna. Bagaimana Keadaan Anda Hari Ini?) Saya Rasa Saya Demam) Anda)Document12 paginiDialog 1: Siang, Anna. Bagaimana Keadaan Anda Hari Ini?) Saya Rasa Saya Demam) Anda)KLINIK ADHYAKSA KEJAKSAAN TINGGI SUMATERA UTARAÎncă nu există evaluări
- Hema 5250Document2 paginiHema 5250GodwinÎncă nu există evaluări
- Cell SignalingDocument1 paginăCell SignalingNathan Stuart The Retarded idiotÎncă nu există evaluări
- Building DNA Student ExplorationDocument5 paginiBuilding DNA Student Explorationaakeelah37Încă nu există evaluări
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDocument7 paginiKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderÎncă nu există evaluări
- Istanbul Jeoloji Harita - CompressedDocument21 paginiIstanbul Jeoloji Harita - Compressedk9chqk26t9Încă nu există evaluări
- Exam Bio-Organic Chemistry (8S140) Thursday November 23, 2008 14.00-17.00 H This Exam Consists of 7 Questions. Explain Your Answers Clearly. Answers May Be Given in English or DutchDocument5 paginiExam Bio-Organic Chemistry (8S140) Thursday November 23, 2008 14.00-17.00 H This Exam Consists of 7 Questions. Explain Your Answers Clearly. Answers May Be Given in English or DutchSergeyÎncă nu există evaluări