S-ar putea să vă placă și
- FDA C19 VaccinesDocument27 paginiFDA C19 VaccinesAll News Pipeline100% (1)
- Osmosis InversaDocument1 paginăOsmosis InversafresitoÎncă nu există evaluări
- Ross General Quote Request Form PDFDocument2 paginiRoss General Quote Request Form PDFfresitoÎncă nu există evaluări
- Black Vipers Windows 10 Service Configurations Black ViperDocument42 paginiBlack Vipers Windows 10 Service Configurations Black ViperfresitoÎncă nu există evaluări
- Ross General BrochureDocument16 paginiRoss General BrochurefresitoÎncă nu există evaluări
- Osmosis InversaDocument1 paginăOsmosis InversafresitoÎncă nu există evaluări
- Brochure E2 ERPDocument13 paginiBrochure E2 ERPfresitoÎncă nu există evaluări
- India VaccineDocument59 paginiIndia VaccinefresitoÎncă nu există evaluări
- Birth: Control ReviewDocument8 paginiBirth: Control ReviewfresitoÎncă nu există evaluări
- 4090-9001 Modulo de Monitoreo IDNetDocument4 pagini4090-9001 Modulo de Monitoreo IDNetwilfredo macoÎncă nu există evaluări
- Estimating Sheet Metal Fabrication Costs v3 PDFDocument58 paginiEstimating Sheet Metal Fabrication Costs v3 PDFtassanaiÎncă nu există evaluări
- Comparative Analysis of Process Flow Configurations For Ethanol DehydrationDocument10 paginiComparative Analysis of Process Flow Configurations For Ethanol DehydrationfresitoÎncă nu există evaluări
- Quest SARS IfuDocument18 paginiQuest SARS IfuZsi GaÎncă nu există evaluări
- EL Hospitality 1 UpdatedDocument12 paginiEL Hospitality 1 UpdatedfresitoÎncă nu există evaluări
- I-9105R Intelligent Reflective Beam Detector Issue3.06Document26 paginiI-9105R Intelligent Reflective Beam Detector Issue3.06fresitoÎncă nu există evaluări
- Mixers: S I L V E R S O NDocument16 paginiMixers: S I L V E R S O NfresitoÎncă nu există evaluări
- The Confidence Factor: Personal AssessmentDocument12 paginiThe Confidence Factor: Personal AssessmentGhulam MustafaÎncă nu există evaluări
- Elec A 02Document1 paginăElec A 02fresitoÎncă nu există evaluări
- Assignment 2 - Plane Stress State and Principal Stresses: Deformed Configuration Reference ConfigurationDocument1 paginăAssignment 2 - Plane Stress State and Principal Stresses: Deformed Configuration Reference ConfigurationfresitoÎncă nu există evaluări
- ASHRAE 90.1 Prescriptive Wall Insulation RequirementsDocument2 paginiASHRAE 90.1 Prescriptive Wall Insulation RequirementsfresitoÎncă nu există evaluări
- EnergySaver Factory Assembled in Plane Rooflights BrochureDocument12 paginiEnergySaver Factory Assembled in Plane Rooflights BrochurefresitoÎncă nu există evaluări
- Problems at The Cumene Production Facility (Sim) PDFDocument17 paginiProblems at The Cumene Production Facility (Sim) PDFfresitoÎncă nu există evaluări
- Elec A 01Document1 paginăElec A 01fresitoÎncă nu există evaluări
- 2015-Tackling Difficult Mixing ProblemsDocument8 pagini2015-Tackling Difficult Mixing Problemsjuan sepulvedaÎncă nu există evaluări
- 1263 3507 1 PB PDFDocument5 pagini1263 3507 1 PB PDFfresitoÎncă nu există evaluări
- Building A Home Distillation ApparatusDocument78 paginiBuilding A Home Distillation Apparatusres06suc100% (7)
- Distillation HandbookDocument94 paginiDistillation HandbookEr Bali Pandhare100% (2)
- The Lie Behind The Lie DetectorDocument220 paginiThe Lie Behind The Lie DetectorAntiPolygraph.org95% (22)
- Making Gin & Vodka A Professional Guide For Amateur DistillersDocument91 paginiMaking Gin & Vodka A Professional Guide For Amateur Distillersbigredmutt88% (8)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (120)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Transfer To Definitive Care EcaDocument40 paginiTransfer To Definitive Care EcaFahrurido Kusbari100% (1)
- The Independence of Combinatory Semantic Processing: Evidence From Event-Related PotentialsDocument21 paginiThe Independence of Combinatory Semantic Processing: Evidence From Event-Related Potentialsdkm2030Încă nu există evaluări
- Biology Neet Question - EnglishDocument377 paginiBiology Neet Question - Englishjoyjebarani0% (1)
- Week 5: Module 9:central Nervous System and Special Senses Time Table (2nd Year Medicine, Semester 3: 2011-2012)Document17 paginiWeek 5: Module 9:central Nervous System and Special Senses Time Table (2nd Year Medicine, Semester 3: 2011-2012)Hairul AnuarÎncă nu există evaluări
- Extroverts (Also Spelled Extraverts: ExtroversionDocument8 paginiExtroverts (Also Spelled Extraverts: Extroversionrams4u_001Încă nu există evaluări
- Assignment Module 3Document14 paginiAssignment Module 3Asshi Sha50% (4)
- BMJ - Hypovolemic ShockDocument5 paginiBMJ - Hypovolemic ShockSamer Darwiche Yasiin100% (1)
- SRIVASTAVA & SINGH, 2008 - Brissid Echinoid Eupatagus From The Khuiala Formation, Jaisalmer District, Rajasthan, IndiaDocument6 paginiSRIVASTAVA & SINGH, 2008 - Brissid Echinoid Eupatagus From The Khuiala Formation, Jaisalmer District, Rajasthan, Indiachindas4321Încă nu există evaluări
- Non-Pharmacological Treatment Options inDocument9 paginiNon-Pharmacological Treatment Options inkemal ghazaliÎncă nu există evaluări
- 3.14.15 Ymok Painmgmt LN ImagesDocument56 pagini3.14.15 Ymok Painmgmt LN ImagesSandar bandÎncă nu există evaluări
- Floating Leaf Disk Assay For Apes Final 2019 AprilDocument11 paginiFloating Leaf Disk Assay For Apes Final 2019 AprilJohn OsborneÎncă nu există evaluări
- FITT 1 Updated HandoutsDocument25 paginiFITT 1 Updated HandoutsJolina VillalobosÎncă nu există evaluări
- Steps HemodialysisDocument3 paginiSteps HemodialysisCes LoÎncă nu există evaluări
- Mumbai AyurvedaDocument2 paginiMumbai AyurvedaAditya SinghÎncă nu există evaluări
- ABC Model and Floral Evolution: ReviewDocument7 paginiABC Model and Floral Evolution: ReviewHairunisa FebrianiÎncă nu există evaluări
- Answer Key P2Document5 paginiAnswer Key P2Marie Stephan De Gracia TanÎncă nu există evaluări
- Immunology Dental Caries PDFDocument29 paginiImmunology Dental Caries PDFTio AjhaÎncă nu există evaluări
- Rula SheetDocument2 paginiRula SheetCarl Christian CabelloÎncă nu există evaluări
- Electromyography I PDFDocument13 paginiElectromyography I PDFkeithtrahÎncă nu există evaluări
- Module 1. General and Special Questions of Clinical Laboratory Diagnostics Text Test QuestionDocument229 paginiModule 1. General and Special Questions of Clinical Laboratory Diagnostics Text Test QuestionA.h.MuradÎncă nu există evaluări
- Pacemaker Invasive Cardiac PacingDocument57 paginiPacemaker Invasive Cardiac PacingAhmad Khalil Ahmad Al-SadiÎncă nu există evaluări
- AbdomenDocument10 paginiAbdomenironÎncă nu există evaluări
- TranslationDocument1 paginăTranslationGianne AbcedeÎncă nu există evaluări
- WBI11 01 Que 20220108Document28 paginiWBI11 01 Que 20220108Hammodshan 7Încă nu există evaluări
- Pathophysiology of Biliary-Type Abdominal Pain: Bertha Van Der DijsDocument3 paginiPathophysiology of Biliary-Type Abdominal Pain: Bertha Van Der DijsDina RuslinaÎncă nu există evaluări
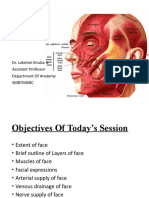
- Dr. Lakshmi Kiruba N Assistant Professor Department of Anatomy SmbtimsrcDocument25 paginiDr. Lakshmi Kiruba N Assistant Professor Department of Anatomy Smbtimsrclakshmi kirubaÎncă nu există evaluări
- Renal Physiology PDFDocument12 paginiRenal Physiology PDFLolitaÎncă nu există evaluări
- Unit 5.: Facultad de Medicina Humana "Daniel Alcides Carrión Garcia"Document12 paginiUnit 5.: Facultad de Medicina Humana "Daniel Alcides Carrión Garcia"MARCOABADÎncă nu există evaluări
- Anaesthetic Implications in A Case of Indian Patient of Xeroderma PigmentosumDocument5 paginiAnaesthetic Implications in A Case of Indian Patient of Xeroderma PigmentosumSwapnil AswarÎncă nu există evaluări
- Assessing Thorax and LungsDocument46 paginiAssessing Thorax and LungsmyfilesÎncă nu există evaluări