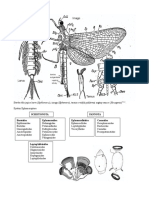
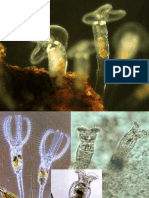
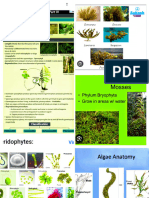
S-ar putea să vă placă și
- Chlo Rophy TaDocument12 paginiChlo Rophy TaRaul Eduardo Beltran CabellÎncă nu există evaluări
- The Princeton Field Guide to Prehistoric MammalsDe la EverandThe Princeton Field Guide to Prehistoric MammalsEvaluare: 5 din 5 stele5/5 (5)
- ChlorophytaDocument12 paginiChlorophytaErick Oriol Sanchez Vargas0% (1)
- Coral Genera of WIO-2015Document57 paginiCoral Genera of WIO-2015Frankie AndrewsÎncă nu există evaluări
- ANGELIKA JOY LOZANO - BS BIO 1-2 - Activity 3 Fish Systematics Laboratory ActivityDocument17 paginiANGELIKA JOY LOZANO - BS BIO 1-2 - Activity 3 Fish Systematics Laboratory ActivityAngelika Joy LozanoÎncă nu există evaluări
- Mode of Reproduction: Invertebrate Animals: October 2018Document11 paginiMode of Reproduction: Invertebrate Animals: October 2018Sophia Pauline HernandezÎncă nu există evaluări
- Introduction To The Endoparasites: NematodesDocument24 paginiIntroduction To The Endoparasites: NematodesdanielaÎncă nu există evaluări
- Freshwater Turtles & Terrapins DemoDocument11 paginiFreshwater Turtles & Terrapins DemoSonny ToshiroÎncă nu există evaluări
- World of KongDocument68 paginiWorld of KongVaszkó Zsolt100% (1)
- ANGELIKA JOY LOZANO - BS BIO 1-2 - Activity 3 Fish Systematics Laboratory Activity PDFDocument17 paginiANGELIKA JOY LOZANO - BS BIO 1-2 - Activity 3 Fish Systematics Laboratory Activity PDFAngelika Joy LozanoÎncă nu există evaluări
- Relatorio - Anual - 2020 - PlantelDocument9 paginiRelatorio - Anual - 2020 - PlantelRaphaël RoisÎncă nu există evaluări
- 03 DinosauriaDocument32 pagini03 DinosauriaMadelyn MackintoshÎncă nu există evaluări
- Batoidea: Temporal Range: Early Triassic-RecentDocument7 paginiBatoidea: Temporal Range: Early Triassic-RecentNirmal BhowmickÎncă nu există evaluări
- Cas Common Prohibited ReptilesDocument33 paginiCas Common Prohibited Reptilesdany alarconÎncă nu există evaluări
- 13.b IntertebratesDocument106 pagini13.b Intertebrateschristinejoy.pineda44Încă nu există evaluări
- Echinodermata TaxaDocument3 paginiEchinodermata TaxaDave RapaconÎncă nu există evaluări
- AlveolateDocument8 paginiAlveolatex456456456xÎncă nu există evaluări
- Schistonota Baetoidea Ephemeroidea Pannota Ephemerelloidea CaenoideaDocument6 paginiSchistonota Baetoidea Ephemeroidea Pannota Ephemerelloidea CaenoideaTomáš ŠpačekÎncă nu există evaluări
- Famili Ikan HiasDocument13 paginiFamili Ikan HiasIvanN2NÎncă nu există evaluări
- Adkins W.S. 1928 - Handbook of Texas Cretaceous FossilsDocument383 paginiAdkins W.S. 1928 - Handbook of Texas Cretaceous FossilsNicolleau PhilippeÎncă nu există evaluări
- 108 Lecture 1 Spring 2023Document22 pagini108 Lecture 1 Spring 2023Chevy WhiteÎncă nu există evaluări
- 07 NeornithischiaDocument38 pagini07 NeornithischiaMadelyn MackintoshÎncă nu există evaluări
- Chalcidoptera Emissalis - WikipediaDocument2 paginiChalcidoptera Emissalis - WikipediaRodrigo1984Încă nu există evaluări
- Bio25: Vertebrate Structure & Function: Janice V. NG, MSC Department of Biology, Cas, Upm Sy 2015-2016Document43 paginiBio25: Vertebrate Structure & Function: Janice V. NG, MSC Department of Biology, Cas, Upm Sy 2015-2016Ruby GoÎncă nu există evaluări
- Phylum Subphylum Class Subclass Superorder Order Suborder: Acanthodii PlacodermiDocument14 paginiPhylum Subphylum Class Subclass Superorder Order Suborder: Acanthodii PlacodermiDimple May Gianne DumaguitÎncă nu există evaluări
- l7 Rotifera 07Document17 paginil7 Rotifera 07Femil adellaÎncă nu există evaluări
- Subclasa Ordin Familia SpeciaDocument5 paginiSubclasa Ordin Familia SpeciaClaudinia CazacÎncă nu există evaluări
- Evolutionary History, Molecular Systematics and Evolutionary Ecology of Canidae (Wang Et Al., 2006) PDFDocument18 paginiEvolutionary History, Molecular Systematics and Evolutionary Ecology of Canidae (Wang Et Al., 2006) PDFFrancisco J. OvalleÎncă nu există evaluări
- Phylogeny and The Tree of Life: BCOR 012 February 4,7, 2011Document36 paginiPhylogeny and The Tree of Life: BCOR 012 February 4,7, 2011Jhanray DoromalÎncă nu există evaluări
- Zooplankton Guide 27Document272 paginiZooplankton Guide 27Ratna AvitasariÎncă nu există evaluări
- The Cichlids Yearbook Vol 1Document22 paginiThe Cichlids Yearbook Vol 1elisav100% (1)
- The Classification of FishesDocument27 paginiThe Classification of FishesBhuvneshwar Gauro100% (1)
- AlgaeDocument7 paginiAlgaeRavi NagraÎncă nu există evaluări
- Entomolgy Chapter 6 - Evolution and SystematicsDocument28 paginiEntomolgy Chapter 6 - Evolution and SystematicsTsega zeab KebedeÎncă nu există evaluări
- Parasitology Chart 211Document20 paginiParasitology Chart 211Sanjay Kumar100% (1)
- Birs Panama - 1965 Vol 150 PT 1Document488 paginiBirs Panama - 1965 Vol 150 PT 1Alejandra ArreolaÎncă nu există evaluări
- Ipers: Iste DES Serpents Venimeux DU MondeDocument4 paginiIpers: Iste DES Serpents Venimeux DU MondeSimon LoupasÎncă nu există evaluări
- The Origins of Life On Earth: Biology 1Document30 paginiThe Origins of Life On Earth: Biology 1Margareth RiveraÎncă nu există evaluări
- Acartia Sp. Adultes Acartia Sp. Adultes Acartia Sp. CopepoditDocument21 paginiAcartia Sp. Adultes Acartia Sp. Adultes Acartia Sp. CopepoditErick Oriol Sanchez VargasÎncă nu există evaluări
- Stuttgarter-Beitraege-Naturkunde 357 A 0001-0055 PDFDocument55 paginiStuttgarter-Beitraege-Naturkunde 357 A 0001-0055 PDFJOSE FRANCISCO FRANCO NAVIAÎncă nu există evaluări
- Bats Are Chiroptera : Eocene PresentDocument23 paginiBats Are Chiroptera : Eocene PresentrachuÎncă nu există evaluări
- Plant ClassificationDocument3 paginiPlant ClassificationAshutosh MishraÎncă nu există evaluări
- Castorimorpha - WikipediaDocument2 paginiCastorimorpha - Wikipediascribd45554Încă nu există evaluări
- Chmiel Ews Kik Raye Sky PreviewDocument16 paginiChmiel Ews Kik Raye Sky PreviewSHEEN ALUBAÎncă nu există evaluări
- Laporan Praktikum Parasitologi Iii: "Klasifikasi Arthropoda"Document6 paginiLaporan Praktikum Parasitologi Iii: "Klasifikasi Arthropoda"Winda NashirahÎncă nu există evaluări
- OiuyuyuyDocument7 paginiOiuyuyuyJessica Asitimbay ZuritaÎncă nu există evaluări
- Platyhelminthes - 2nd Class PDFDocument18 paginiPlatyhelminthes - 2nd Class PDFAbdulla Hil KafiÎncă nu există evaluări
- Pollen Flora of Pakistan-XLVII EuphorbiaceaeDocument13 paginiPollen Flora of Pakistan-XLVII EuphorbiaceaeBrenda Leidy Orosco ArangoÎncă nu există evaluări
- Pathology of Birds - An Overview: H. L. ShivaprasadDocument47 paginiPathology of Birds - An Overview: H. L. ShivaprasadDavin ChioÎncă nu există evaluări
- ParasiteofsnakesinThailand PDFDocument13 paginiParasiteofsnakesinThailand PDFEstefania MoralesÎncă nu există evaluări
- Helminths 12Document34 paginiHelminths 12malakaiad212Încă nu există evaluări
- Bird List TambopataDocument12 paginiBird List TambopataYuliana GomezÎncă nu există evaluări
- ICAR Animal Science Short NotesDocument252 paginiICAR Animal Science Short NotesDr AMEER SUHAIL LOYOLITEÎncă nu există evaluări
- CoralsDocument2 paginiCoralsRyan LonÎncă nu există evaluări
- Wrestling With Crustacean Taxonomy I Can'T Remember What Class I'M in Subphylum CrustaceaDocument10 paginiWrestling With Crustacean Taxonomy I Can'T Remember What Class I'M in Subphylum CrustaceawateereffectÎncă nu există evaluări
- Alula Fauna 2021Document3 paginiAlula Fauna 2021Mohammed Ali QaziÎncă nu există evaluări
- Yasa Maritime VocabularyDocument9 paginiYasa Maritime VocabularySamuel EkpoÎncă nu există evaluări
- A Guide To Orthoptera and Allied Insects: ITO Gen and Homathevi RahmanDocument8 paginiA Guide To Orthoptera and Allied Insects: ITO Gen and Homathevi RahmanRolando CoronelÎncă nu există evaluări
- Kab/ Situs Kode Wilayah Jenis Temuan: Kotak KTK 1/ MMKDocument50 paginiKab/ Situs Kode Wilayah Jenis Temuan: Kotak KTK 1/ MMKMaria MayorÎncă nu există evaluări
- Mammalian Teeth 03 Schulz KornasDocument10 paginiMammalian Teeth 03 Schulz KornasBogdan HaiducÎncă nu există evaluări
- Sheffield Edge Tool MakersDocument48 paginiSheffield Edge Tool MakersnikkicluÎncă nu există evaluări
- European Neogene Mammal ChronologyDocument9 paginiEuropean Neogene Mammal ChronologyBogdan HaiducÎncă nu există evaluări
- Cifelli Ed 1996Document41 paginiCifelli Ed 1996Bogdan HaiducÎncă nu există evaluări
- Haiduc Et Al. 2018Document8 paginiHaiduc Et Al. 2018Bogdan HaiducÎncă nu există evaluări
- The History of Mesowear - A Review, Ackermans 2020Document17 paginiThe History of Mesowear - A Review, Ackermans 2020Bogdan HaiducÎncă nu există evaluări
- Pages From Ward Et Al (2000) Science - EquatoriusDocument1 paginăPages From Ward Et Al (2000) Science - EquatoriusBogdan HaiducÎncă nu există evaluări
- Paleoenvironment in East Java During The Last 25,000 Years As Inferred From Bovid and Cervid Dental Wear Analyses - 2016 Rivals Et AlDocument12 paginiPaleoenvironment in East Java During The Last 25,000 Years As Inferred From Bovid and Cervid Dental Wear Analyses - 2016 Rivals Et AlBogdan HaiducÎncă nu există evaluări
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Document10 paginiTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducÎncă nu există evaluări
- Gr. Cobalcescu Annual Scientific Symposium - en 2019Document2 paginiGr. Cobalcescu Annual Scientific Symposium - en 2019Bogdan HaiducÎncă nu există evaluări
- Dental Microwear Texture Analysis: Technical ConsiderationsDocument11 paginiDental Microwear Texture Analysis: Technical ConsiderationsBogdan HaiducÎncă nu există evaluări
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Document10 paginiTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducÎncă nu există evaluări
- Kelley & Gao (2012) PNAS - Yuanmou Cranium PDFDocument4 paginiKelley & Gao (2012) PNAS - Yuanmou Cranium PDFBogdan HaiducÎncă nu există evaluări
- PastmanualDocument205 paginiPastmanualSheetal BhosaleÎncă nu există evaluări
- Kelley & Gao (2012) PNAS (SI) - Yuanmou CraniumDocument3 paginiKelley & Gao (2012) PNAS (SI) - Yuanmou CraniumBogdan HaiducÎncă nu există evaluări
- Scott Et Al (2009) AJPA - Ouranopithecus Sexual DimorphismDocument12 paginiScott Et Al (2009) AJPA - Ouranopithecus Sexual DimorphismBogdan HaiducÎncă nu există evaluări
- How To Install R PDFDocument2 paginiHow To Install R PDFBogdan HaiducÎncă nu există evaluări
- Hershkovitz Et Al (1997) JHE - Sivapithecus Oral BacteriaDocument6 paginiHershkovitz Et Al (1997) JHE - Sivapithecus Oral BacteriaBogdan HaiducÎncă nu există evaluări
- Anders & Koenigswald (2013) Mammuthus AgesDocument16 paginiAnders & Koenigswald (2013) Mammuthus AgesBogdan HaiducÎncă nu există evaluări
- Why Does Micromorphology Have Such Little Traction in Geoarchaeology - Aldeias Et Al 2016Document11 paginiWhy Does Micromorphology Have Such Little Traction in Geoarchaeology - Aldeias Et Al 2016Bogdan HaiducÎncă nu există evaluări
- Kelley & Alpagut (1999) JHE - Pasalar CaninesDocument7 paginiKelley & Alpagut (1999) JHE - Pasalar CaninesBogdan HaiducÎncă nu există evaluări
- Phytoliths - A Comprehensive Guide For Archaeologists and PaleoecologistsDocument2 paginiPhytoliths - A Comprehensive Guide For Archaeologists and PaleoecologistsBogdan HaiducÎncă nu există evaluări
- 2016 Impact Factor (JCR)Document210 pagini2016 Impact Factor (JCR)dipsiiiÎncă nu există evaluări
- Comparison of Switching Strategies For An Ultrasonic Cleaner PDFDocument5 paginiComparison of Switching Strategies For An Ultrasonic Cleaner PDFBogdan HaiducÎncă nu există evaluări
- Kelley & Etler (1989) AJP - Lufeng Species NumberDocument20 paginiKelley & Etler (1989) AJP - Lufeng Species NumberBogdan HaiducÎncă nu există evaluări
- Evolution and Palaeobiology of PterosaursDocument2 paginiEvolution and Palaeobiology of PterosaursBogdan HaiducÎncă nu există evaluări
- Registration Form SPR 2017Document1 paginăRegistration Form SPR 2017Bogdan HaiducÎncă nu există evaluări
- Baigusheva Et Al - Tooth of Early Generations - QI - 2016Document14 paginiBaigusheva Et Al - Tooth of Early Generations - QI - 2016Bogdan HaiducÎncă nu există evaluări
- First Cenozoic BirdDocument8 paginiFirst Cenozoic BirdBogdan HaiducÎncă nu există evaluări
- CONTENT COURSE W AnsDocument10 paginiCONTENT COURSE W AnsArjay JacobÎncă nu există evaluări
- Geotechnical Report N018 Rev0Document18 paginiGeotechnical Report N018 Rev0Fabian Herrera TroncosoÎncă nu există evaluări
- Multilateral Wells in Production of Thin Oil Reservoirs DissertationDocument5 paginiMultilateral Wells in Production of Thin Oil Reservoirs DissertationFasakin AkinwoleÎncă nu există evaluări
- Amphiboles PDFDocument3 paginiAmphiboles PDFMaria Martha SarmientoÎncă nu există evaluări
- What Is Bauxite?: Mineral RockDocument13 paginiWhat Is Bauxite?: Mineral RockRenesha100% (1)
- Sea Water Intrusion in GreeceDocument13 paginiSea Water Intrusion in GreeceAnonymous BVbpSEÎncă nu există evaluări
- 04 - NTH - Project Report 1-94 Hard Rock Tunnel Boring PDFDocument165 pagini04 - NTH - Project Report 1-94 Hard Rock Tunnel Boring PDFWaqas Sadiq100% (2)
- Mid-Ocean Ridge: People Also AskDocument3 paginiMid-Ocean Ridge: People Also AskBlessing NgonidzasheÎncă nu există evaluări
- Balakrishnan1999 PDFDocument11 paginiBalakrishnan1999 PDFATISH KUMAR DASÎncă nu există evaluări
- Magnetic Interpretation in Three Dimensions Using Euler DeconvolutionDocument12 paginiMagnetic Interpretation in Three Dimensions Using Euler DeconvolutionMutiara IslamiaÎncă nu există evaluări
- Leach Et Al. 2010 PDFDocument33 paginiLeach Et Al. 2010 PDFJulio RamiroÎncă nu există evaluări
- Well Test Interpretation - ENSPM - 1a PDFDocument60 paginiWell Test Interpretation - ENSPM - 1a PDFMygroup 5544100% (1)
- ConclusionDocument3 paginiConclusionPuspita Purnama Sari100% (1)
- Management of Tailings and Waste Rock in Mining ActvitiesDocument557 paginiManagement of Tailings and Waste Rock in Mining ActvitiesPaula TrindadeÎncă nu există evaluări
- Gulf of Mexico Oil Gusher FissureDocument140 paginiGulf of Mexico Oil Gusher FissureVincent J. CataldiÎncă nu există evaluări
- Prehistory of The PhilippinesDocument29 paginiPrehistory of The PhilippinesmhyckÎncă nu există evaluări
- Geography Mock TestDocument20 paginiGeography Mock TestPavan K. TiwariÎncă nu există evaluări
- DR Björn Kjerfve 2016 AUSDocument23 paginiDR Björn Kjerfve 2016 AUSAnna WilliamsÎncă nu există evaluări
- Null 2Document73 paginiNull 2Zoey EliseÎncă nu există evaluări
- Sedex Poster Sharanya VaratharajooDocument1 paginăSedex Poster Sharanya Varatharajooapi-439238002Încă nu există evaluări
- Introduction Resposibilities and AdministrationDocument22 paginiIntroduction Resposibilities and Administrationrdos14Încă nu există evaluări
- Some Evaporite Deposits of India: Proc Indian Natn Sci Acad 78 No. 3 September 2012 Pp. 401-406 Printed in IndiaDocument6 paginiSome Evaporite Deposits of India: Proc Indian Natn Sci Acad 78 No. 3 September 2012 Pp. 401-406 Printed in Indiavaishu2488Încă nu există evaluări
- Biology 405 Organic Evolution Lab ManualDocument9 paginiBiology 405 Organic Evolution Lab ManualZielonaZabaÎncă nu există evaluări
- 1 Basic Concepts PDFDocument4 pagini1 Basic Concepts PDFHummayoun KhanÎncă nu există evaluări
- SCTM 15Document38 paginiSCTM 15Zhi Ming CheahÎncă nu există evaluări
- Lecture6 Locating EpicenterDocument15 paginiLecture6 Locating EpicenterAndrea Sochayseng SolijonÎncă nu există evaluări
- Why Re-FracDocument45 paginiWhy Re-Fracrobertobg1Încă nu există evaluări
- Fracture Characterization and Modelling of A Tight Carbonate ReservoirDocument16 paginiFracture Characterization and Modelling of A Tight Carbonate ReservoirAhmad AdvinÎncă nu există evaluări
- Allen, Et Al., (2015)Document24 paginiAllen, Et Al., (2015)chris17tianÎncă nu există evaluări
- Carbonates and Matrix AcidizingDocument37 paginiCarbonates and Matrix AcidizingPhintulBingungÎncă nu există evaluări
- Merle's Door: Lessons from a Freethinking DogDe la EverandMerle's Door: Lessons from a Freethinking DogEvaluare: 4 din 5 stele4/5 (326)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueDe la EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueEvaluare: 5 din 5 stele5/5 (3)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueDe la EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueEvaluare: 4.5 din 5 stele4.5/5 (26)
- The Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationDe la EverandThe Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationEvaluare: 4 din 5 stele4/5 (37)
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessDe la EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessÎncă nu există evaluări
- Will's Red Coat: The Story of One Old Dog Who Chose to Live AgainDe la EverandWill's Red Coat: The Story of One Old Dog Who Chose to Live AgainEvaluare: 4.5 din 5 stele4.5/5 (18)
- Arthur: The Dog Who Crossed the Jungle to Find a HomeDe la EverandArthur: The Dog Who Crossed the Jungle to Find a HomeEvaluare: 4.5 din 5 stele4.5/5 (20)
- Show Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredDe la EverandShow Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredEvaluare: 3.5 din 5 stele3.5/5 (13)
- The Other End of the Leash: Why We Do What We Do Around DogsDe la EverandThe Other End of the Leash: Why We Do What We Do Around DogsEvaluare: 5 din 5 stele5/5 (65)
- Come Back, Como: Winning the Heart of a Reluctant DogDe la EverandCome Back, Como: Winning the Heart of a Reluctant DogEvaluare: 3.5 din 5 stele3.5/5 (10)
- An Eagle Named Freedom: My True Story of a Remarkable FriendshipDe la EverandAn Eagle Named Freedom: My True Story of a Remarkable FriendshipÎncă nu există evaluări
- Dogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowDe la EverandDogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowÎncă nu există evaluări
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsDe la EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsÎncă nu există evaluări
- Inside of a Dog: What Dogs See, Smell, and KnowDe la EverandInside of a Dog: What Dogs See, Smell, and KnowEvaluare: 4 din 5 stele4/5 (390)
- Your Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesDe la EverandYour Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesEvaluare: 4 din 5 stele4/5 (31)
- How Dogs Think: What the World Looks Like to Them and Why They Act the Way They DoDe la EverandHow Dogs Think: What the World Looks Like to Them and Why They Act the Way They DoEvaluare: 4 din 5 stele4/5 (46)
- Lucky Dog Lessons: Train Your Dog in 7 DaysDe la EverandLucky Dog Lessons: Train Your Dog in 7 DaysEvaluare: 4.5 din 5 stele4.5/5 (41)
- Second Chances: A Marine, His Dog, and Finding RedemptionDe la EverandSecond Chances: A Marine, His Dog, and Finding RedemptionÎncă nu există evaluări
- Edward's Menagerie: Dogs: 50 canine crochet patternsDe la EverandEdward's Menagerie: Dogs: 50 canine crochet patternsEvaluare: 3 din 5 stele3/5 (5)
- The Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenDe la EverandThe Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenEvaluare: 4.5 din 5 stele4.5/5 (49)
- Our Dogs, Ourselves: The Story of a Singular BondDe la EverandOur Dogs, Ourselves: The Story of a Singular BondEvaluare: 4 din 5 stele4/5 (21)
- Puppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!De la EverandPuppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!Evaluare: 5 din 5 stele5/5 (85)
- Julia's Cats: Julia Child's Life in the Company of CatsDe la EverandJulia's Cats: Julia Child's Life in the Company of CatsEvaluare: 4.5 din 5 stele4.5/5 (24)
- Decoding Your Cat: The Ultimate Experts Explain Common Cat Behaviors and Reveal How to Prevent or Change Unwanted OnesDe la EverandDecoding Your Cat: The Ultimate Experts Explain Common Cat Behaviors and Reveal How to Prevent or Change Unwanted OnesEvaluare: 4 din 5 stele4/5 (6)
- How to Be a Good Creature: A Memoir in Thirteen AnimalsDe la EverandHow to Be a Good Creature: A Memoir in Thirteen AnimalsEvaluare: 4 din 5 stele4/5 (223)