S-ar putea să vă placă și
- Fishes in Lagoons and Estuaries in the Mediterranean 1: Diversity, Bioecology and ExploitationDe la EverandFishes in Lagoons and Estuaries in the Mediterranean 1: Diversity, Bioecology and ExploitationÎncă nu există evaluări
- Biology of Benthic Organisms: 11th European Symposium on Marine Biology, Galway, October 1976De la EverandBiology of Benthic Organisms: 11th European Symposium on Marine Biology, Galway, October 1976B. F. KeeganEvaluare: 5 din 5 stele5/5 (1)
- Montero-Pau - 2008 - OCEANOGRAPHY METHODS Application of An Inexpensive and High-Throughput Genomic DNA Extraction Method For The MolecDocument5 paginiMontero-Pau - 2008 - OCEANOGRAPHY METHODS Application of An Inexpensive and High-Throughput Genomic DNA Extraction Method For The MolecFlavio DávilaÎncă nu există evaluări
- J Aquaculture 2018 11 042Document29 paginiJ Aquaculture 2018 11 042Miguel García PortaÎncă nu există evaluări
- Benetti 2010 Growth Rates of Cobia (Rachycentron Canadum) Cultured in Open Ocean Submerged Cages in The CaribbeanDocument7 paginiBenetti 2010 Growth Rates of Cobia (Rachycentron Canadum) Cultured in Open Ocean Submerged Cages in The CaribbeanThais BatistaÎncă nu există evaluări
- Experimental Rearing of The Sea Urchinparacentrotus Lividus Fed With Discards of The Lettucelactuca Sativain A Sea-Based SystemDocument6 paginiExperimental Rearing of The Sea Urchinparacentrotus Lividus Fed With Discards of The Lettucelactuca Sativain A Sea-Based SystemMorris MorrisÎncă nu există evaluări
- F060 01 1412 2015 ObjectivesDocument11 paginiF060 01 1412 2015 ObjectivesIan FredrickÎncă nu există evaluări
- Effects of Stocking Density On Survival, Growth andDocument15 paginiEffects of Stocking Density On Survival, Growth andyesim celikÎncă nu există evaluări
- Journal of Experimental Marine Biology and Ecology: Darlan J.B. Simith, Karen DieleDocument7 paginiJournal of Experimental Marine Biology and Ecology: Darlan J.B. Simith, Karen DieleharisankarhsÎncă nu există evaluări
- Effects of Salinity and Temperature On The Growth and Survival of New Zealand Shortfin, Anguilla Australis, and Longfin, ADocument9 paginiEffects of Salinity and Temperature On The Growth and Survival of New Zealand Shortfin, Anguilla Australis, and Longfin, AMarianus LeinÎncă nu există evaluări
- Aquaculture Potential of The Common OctopusDocument18 paginiAquaculture Potential of The Common OctopusDoctorEmirÎncă nu există evaluări
- 1 s2.0 S0044848604006052 Main PDFDocument10 pagini1 s2.0 S0044848604006052 Main PDFVíctor GuardiolaÎncă nu există evaluări
- Modelling The Growth of Crustacean Species: Reviews in Fish Biology and Fisheries March 2011Document32 paginiModelling The Growth of Crustacean Species: Reviews in Fish Biology and Fisheries March 2011Juhenzie PindanÎncă nu există evaluări
- In Uence of Modified Atmosphere Packaging On The Chilled Shelf Life of Gutted Farmed Bass (Dicentrarchus Labrax)Document9 paginiIn Uence of Modified Atmosphere Packaging On The Chilled Shelf Life of Gutted Farmed Bass (Dicentrarchus Labrax)pattysaborio520Încă nu există evaluări
- Biology of Deep-Water Chondrichthyans Introduction. Deep Sea Research Part II Topical Studies in Oceanography, 115, 1-10Document10 paginiBiology of Deep-Water Chondrichthyans Introduction. Deep Sea Research Part II Topical Studies in Oceanography, 115, 1-10Alfredo Sotelo'sÎncă nu există evaluări
- Arnaud, L 2003Document10 paginiArnaud, L 2003Jorge MartínezÎncă nu există evaluări
- Polyculture of Pacific White Shrimp Litopenaeus Vannamei Giant 1Document6 paginiPolyculture of Pacific White Shrimp Litopenaeus Vannamei Giant 1SteveTillalsoÎncă nu există evaluări
- The Effects of Stocking Density On Key Biological Parameters Influencing Culture Productivity of The Cananoid Copepod Parvocalanus CrassirostrisDocument7 paginiThe Effects of Stocking Density On Key Biological Parameters Influencing Culture Productivity of The Cananoid Copepod Parvocalanus CrassirostrisJordan IsmaelÎncă nu există evaluări
- Efectos Del CO 2 Elevado en La Reproducción de Dos Copépodos CalanoideosDocument7 paginiEfectos Del CO 2 Elevado en La Reproducción de Dos Copépodos CalanoideosISABEL SANCHEZ RIVERAÎncă nu există evaluări
- Marine Journal MaritimeDocument7 paginiMarine Journal MaritimeSyahrul Rizki RamadhanÎncă nu există evaluări
- Aquaculture: E. Paredes, J. Bellas, D. CostasDocument4 paginiAquaculture: E. Paredes, J. Bellas, D. CostasCamila OrtizÎncă nu există evaluări
- Effect of Vacuum and Modified Atmosphere Packaging On The Microbiological, Chemical and Sensory Properties of Tropical Red Drum (Sciaenops Ocellatus) Fillets Stored at 4 °CDocument49 paginiEffect of Vacuum and Modified Atmosphere Packaging On The Microbiological, Chemical and Sensory Properties of Tropical Red Drum (Sciaenops Ocellatus) Fillets Stored at 4 °Cpattysaborio520Încă nu există evaluări
- Polleetal2008Dunaliella PDFDocument8 paginiPolleetal2008Dunaliella PDFCarmen Palazon AlbaÎncă nu există evaluări
- Effect of Different Tank Colours On Some Productive Parameters of European CatfishDocument5 paginiEffect of Different Tank Colours On Some Productive Parameters of European Catfishmcoada82Încă nu există evaluări
- (Ferrari Et Al., 2019) PDFDocument6 pagini(Ferrari Et Al., 2019) PDFhre eyhehrhhÎncă nu există evaluări
- Stable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodDocument5 paginiStable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodMarco Antonio SolisÎncă nu există evaluări
- Santos Et Al 2021 (Tetra Olho de Fogo)Document5 paginiSantos Et Al 2021 (Tetra Olho de Fogo)Higo AbeÎncă nu există evaluări
- Cobia BibliographyDocument97 paginiCobia Bibliographynhimdoc2016Încă nu există evaluări
- Advanc 2Document17 paginiAdvanc 2john pierreÎncă nu există evaluări
- Holste Peck 2006Document11 paginiHolste Peck 2006Miguel Ángel Gallego DomínguezÎncă nu există evaluări
- Development of New Quality Index Method (QIM) Schemes For Cuttlefish (Sepia o Cinalis) and Broadtail Shortfin Squid (Illex Coindetii)Document8 paginiDevelopment of New Quality Index Method (QIM) Schemes For Cuttlefish (Sepia o Cinalis) and Broadtail Shortfin Squid (Illex Coindetii)rifayuzÎncă nu există evaluări
- Use Only: Hard Times For Catadromous Fish: The Case of The European Eel Anguilla Anguilla (L. 1758)Document18 paginiUse Only: Hard Times For Catadromous Fish: The Case of The European Eel Anguilla Anguilla (L. 1758)philosophy-thoughtÎncă nu există evaluări
- Coral Microbiome Diversity Reflects Mass Coral Bleaching Susceptibility During The 2016 El Niño Heat WaveDocument19 paginiCoral Microbiome Diversity Reflects Mass Coral Bleaching Susceptibility During The 2016 El Niño Heat WaveSarah SobhiÎncă nu există evaluări
- Culture Experiments With Oithona Oculata and Its Adventages As Food For Marine Fish LarvaeDocument13 paginiCulture Experiments With Oithona Oculata and Its Adventages As Food For Marine Fish LarvaeJordan IsmaelÎncă nu există evaluări
- Reproductive Biology of The Commercial Sea Cucumber Athyonidium Chilensis (Holothuroidea: Dendrochirotida) In..Document14 paginiReproductive Biology of The Commercial Sea Cucumber Athyonidium Chilensis (Holothuroidea: Dendrochirotida) In..Lopa LopaÎncă nu există evaluări
- Silva Et Al 2021 - Bromatological Mexilhão 2659-14551-1-PBDocument11 paginiSilva Et Al 2021 - Bromatological Mexilhão 2659-14551-1-PBcarloskoaÎncă nu există evaluări
- The Biology and Economics of Coral Growth: Invited ReviewDocument14 paginiThe Biology and Economics of Coral Growth: Invited Reviewluiggi zambranoÎncă nu există evaluări
- Crassostrea GigasDocument10 paginiCrassostrea GigassantiagoÎncă nu există evaluări
- Rodrigues Galdino2009Document14 paginiRodrigues Galdino2009Belay AbdissaÎncă nu există evaluări
- Borell Et AlDocument8 paginiBorell Et Alapi-3828346Încă nu există evaluări
- Produccion Cacao Colombia InteresDocument9 paginiProduccion Cacao Colombia InteresJENNIFER TATIANA PORRAS OLIVEROSÎncă nu există evaluări
- Reproductive Response of Nile Tilapia (Oreochromis Niloticus) To Photoperiodic Manipulation Effects On Spawning Periodicity, Fecundity and Egg SizeDocument16 paginiReproductive Response of Nile Tilapia (Oreochromis Niloticus) To Photoperiodic Manipulation Effects On Spawning Periodicity, Fecundity and Egg SizeDARWIN SALAMANCA CANSAYAÎncă nu există evaluări
- tải xuốngDocument14 paginitải xuốngMr CuongÎncă nu există evaluări
- Balbisiana) As Organic Fertilizer On The Growth and Yield of PechayDocument6 paginiBalbisiana) As Organic Fertilizer On The Growth and Yield of PechayXÏX LXÏXÎncă nu există evaluări
- Potencial de Microalgas Bentónicas Del Mar Caribe Como Alimento en MariculturaDocument13 paginiPotencial de Microalgas Bentónicas Del Mar Caribe Como Alimento en MariculturaMirele BarbosaÎncă nu există evaluări
- TMP 2 CCDocument13 paginiTMP 2 CCFrontiersÎncă nu există evaluări
- Fitoplancton Do Lago Das Garcas Sao Paulo Brasil UDocument30 paginiFitoplancton Do Lago Das Garcas Sao Paulo Brasil UPaulo VieiraÎncă nu există evaluări
- Optimization of Biomass, Total Carotenoids and Astaxanthin Production in Haematococcus Pluvialis Flotow Strain Steptoe (Nevada, USA) Under Laboratory ConditionsDocument15 paginiOptimization of Biomass, Total Carotenoids and Astaxanthin Production in Haematococcus Pluvialis Flotow Strain Steptoe (Nevada, USA) Under Laboratory ConditionsNelson Gonzalez MedelÎncă nu există evaluări
- Seminar ReportDocument21 paginiSeminar ReportRosel Conog BugayÎncă nu există evaluări
- Tafreshi Et Al-2009-Journal of Applied Microbiology PDFDocument22 paginiTafreshi Et Al-2009-Journal of Applied Microbiology PDFAntonMauntÎncă nu există evaluări
- CBP New Method HistopathologyDocument8 paginiCBP New Method HistopathologyRenanda Baghaz PutraÎncă nu există evaluări
- TALABA COLORED CHALK 1 5final 1Document40 paginiTALABA COLORED CHALK 1 5final 1Mhel AmarÎncă nu există evaluări
- 2012 Ecological - ISMEDocument14 pagini2012 Ecological - ISMEVERONICA BLANCOÎncă nu există evaluări
- Food Chemistry: M.D. Hernández, M.B. López, A. Álvarez, E. Ferrandini, B. García García, M.D. GarridoDocument9 paginiFood Chemistry: M.D. Hernández, M.B. López, A. Álvarez, E. Ferrandini, B. García García, M.D. Garridoarif yusuf julionartaÎncă nu există evaluări
- Escherichia Coli Populations in Great Lakes Waterfowl Exhibit SpatialDocument6 paginiEscherichia Coli Populations in Great Lakes Waterfowl Exhibit SpatialMohamed Amin MazenÎncă nu există evaluări
- J. Shellfish Res 2008 C. Corteziensis Growth Gametogenic Activity CIDocument11 paginiJ. Shellfish Res 2008 C. Corteziensis Growth Gametogenic Activity CIRomina AlterioÎncă nu există evaluări
- Camargo Et Al 2004Document10 paginiCamargo Et Al 2004rini susilowatiÎncă nu există evaluări
- Transportation and The Effects of Stocking Density On The Survival and Growth of Nile Tilapia, Oreochromis Niloticus (Linnaeus)Document7 paginiTransportation and The Effects of Stocking Density On The Survival and Growth of Nile Tilapia, Oreochromis Niloticus (Linnaeus)Chern YuanÎncă nu există evaluări
- Microsatellite Polymorphism of Seabass (Dicentrarchus Labrax) Breeders: Natural and Cultured StocksDocument7 paginiMicrosatellite Polymorphism of Seabass (Dicentrarchus Labrax) Breeders: Natural and Cultured StocksBilge KarahanÎncă nu există evaluări
- 3IJEEFUSAPR20193Document10 pagini3IJEEFUSAPR20193TJPRC PublicationsÎncă nu există evaluări
- Digital Dash I/O Adapter Configuration: Connector Pin FunctionsDocument8 paginiDigital Dash I/O Adapter Configuration: Connector Pin FunctionsAfeef Ibn AlbraÎncă nu există evaluări
- Reich 0001 PDFDocument87 paginiReich 0001 PDFYordys Domínguez RodríguezÎncă nu există evaluări
- Program 2019 MTAPTL Annual Convention PDFDocument3 paginiProgram 2019 MTAPTL Annual Convention PDFrichardlfigueroaÎncă nu există evaluări
- Different Strategies in TeachingDocument12 paginiDifferent Strategies in TeachingCris MolinaÎncă nu există evaluări
- 20 Heirs of Alfredo Bautista VS LindoDocument3 pagini20 Heirs of Alfredo Bautista VS LindoJerome LeañoÎncă nu există evaluări
- Name: - Date: - Week/s: - 2 - Topic: Examining Trends and FadsDocument3 paginiName: - Date: - Week/s: - 2 - Topic: Examining Trends and FadsRojelyn Conturno100% (1)
- Chapter 3 ContDocument12 paginiChapter 3 ContVenus Frias-AntonioÎncă nu există evaluări
- Gurdan Saini: From Wikipedia, The Free EncyclopediaDocument6 paginiGurdan Saini: From Wikipedia, The Free EncyclopediaRanjeet SinghÎncă nu există evaluări
- STD 12 Accountancy1Document264 paginiSTD 12 Accountancy1Chetan KhonaÎncă nu există evaluări
- Ef3 Englisch 04 14 08052014Document20 paginiEf3 Englisch 04 14 08052014PiPradoÎncă nu există evaluări
- Kindergarten ArchitectureDocument65 paginiKindergarten ArchitectureAnushka Khatri83% (6)
- Hitachi Zx330 3 Series Hydraulic ExcavatorDocument15 paginiHitachi Zx330 3 Series Hydraulic ExcavatorAgung ArdhanaÎncă nu există evaluări
- Thesis TopicsDocument9 paginiThesis TopicsInayath AliÎncă nu există evaluări
- Milling Machine Maintenance, Safety and OperationDocument3 paginiMilling Machine Maintenance, Safety and OperationPPSwain100% (1)
- Legal Framework On Sexual Violence in Sri Lanka - Policy BriefDocument4 paginiLegal Framework On Sexual Violence in Sri Lanka - Policy BriefwmcsrilankaÎncă nu există evaluări
- PDF 20221114 080033 0000Document2 paginiPDF 20221114 080033 0000John Marithe PutunganÎncă nu există evaluări
- Business Advantage Pers Study Book Intermediate PDFDocument98 paginiBusiness Advantage Pers Study Book Intermediate PDFCool Nigga100% (1)
- INSTRUCTIONS FOR USOL PRIVATE REAPPEAR DE January 2022Document4 paginiINSTRUCTIONS FOR USOL PRIVATE REAPPEAR DE January 2022RaghavÎncă nu există evaluări
- Commentary On The Book of NahumDocument9 paginiCommentary On The Book of NahumRev Dr Jeffry Camm JP, MIEPR, MISOPÎncă nu există evaluări
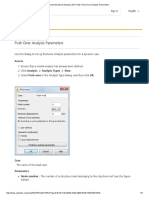
- Robot Structural Analysis 2017 Help - Push Over Analysis ParametersDocument3 paginiRobot Structural Analysis 2017 Help - Push Over Analysis ParametersJustin MusopoleÎncă nu există evaluări
- Q&A FractureDocument13 paginiQ&A FractureRed JimenoÎncă nu există evaluări
- Kids Cooking Teams: That's FreshDocument74 paginiKids Cooking Teams: That's FreshNCB School of Herbalism & Holistic HealthÎncă nu există evaluări
- Zimbabwe National Code Ccorporate GovernanceDocument96 paginiZimbabwe National Code Ccorporate GovernanceHerbert NgwaraiÎncă nu există evaluări
- La Lit Review-Jennifer Draper-2-2Document9 paginiLa Lit Review-Jennifer Draper-2-2api-653567856Încă nu există evaluări
- HCT Baniqued P.D.E. Paper1 Version3 FullpaperDocument8 paginiHCT Baniqued P.D.E. Paper1 Version3 FullpaperJoshua HernandezÎncă nu există evaluări
- Sen Çal KapımıDocument9 paginiSen Çal KapımıMa Gabriela Ojeda CasellaÎncă nu există evaluări
- Onechanbara - Bikini Samurai Squad Game GuideDocument83 paginiOnechanbara - Bikini Samurai Squad Game GuideStefan RoscaÎncă nu există evaluări
- Students List - All SectionsDocument8 paginiStudents List - All SectionsChristian RiveraÎncă nu există evaluări
- Oral ComDocument2 paginiOral ComChristian OwlzÎncă nu există evaluări
- SAGEM FAST 3202 (Livebox) ENGLISHDocument140 paginiSAGEM FAST 3202 (Livebox) ENGLISHspam47spam47Încă nu există evaluări