S-ar putea să vă placă și
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Axillary Node DissectionDocument71 paginiAxillary Node DissectionKartik Kaistha100% (1)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5795)
- DAoC AchievementsDocument13 paginiDAoC AchievementsJoey FieldsÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Survival KitDocument8 paginiSurvival KitedGarKazy77Încă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Skeletal SystemDocument15 paginiSkeletal SystemKarla OñasÎncă nu există evaluări
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Pulmonary TuberculosisDocument4 paginiPulmonary TuberculosisAymen OmerÎncă nu există evaluări
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Ket CambridgeDocument23 paginiKet Cambridgeo100Încă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- PPG Reviewer For ShsDocument5 paginiPPG Reviewer For ShsGoldwin RamirezÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Laboratory Test Normal Range in Us Units Normal Range in Si Units To Convert Ustosi UnitsDocument5 paginiLaboratory Test Normal Range in Us Units Normal Range in Si Units To Convert Ustosi Unitsloloko55113Încă nu există evaluări
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- 007ho13921 - Zeke-P-RedDocument1 pagină007ho13921 - Zeke-P-Redmiguel luzardoÎncă nu există evaluări
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Lista Determinatoare MacronevertebrateDocument2 paginiLista Determinatoare MacronevertebrateramonaÎncă nu există evaluări
- Ingles 5º Unit 3 PDFDocument5 paginiIngles 5º Unit 3 PDFMiguel Angel Llamas DelicadoÎncă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
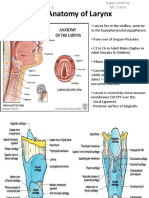
- Anatomy of Larynx - Wai Sheng Xuan Batch 3 1101A11688Document27 paginiAnatomy of Larynx - Wai Sheng Xuan Batch 3 1101A11688Roshan Gill100% (1)
- Coronary Artery Disease - Case StudyDocument19 paginiCoronary Artery Disease - Case StudyJulieteeySarania100% (3)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- 1 SMDocument10 pagini1 SMNurul Hikmah Haetapi AminÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- NPCBB WordDocument6 paginiNPCBB WordNisha Aravindan NairÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- T2 Sains KSSM (Section B &C)Document15 paginiT2 Sains KSSM (Section B &C)Azreen Syazwani Azmi100% (1)
- Chinese Metasoft: Home Products Da Liu Ren Ba Zi Qi Men Dun Jia Feng Shui Ze RiDocument1 paginăChinese Metasoft: Home Products Da Liu Ren Ba Zi Qi Men Dun Jia Feng Shui Ze RiFingiel BluÎncă nu există evaluări
- Tropical Oceans: Word SearchDocument2 paginiTropical Oceans: Word SearchAgrobacterium TumefaciensÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- A Qualitative Research On Pet Owners Using AnimalsDocument16 paginiA Qualitative Research On Pet Owners Using AnimalsJorilou LlamzonÎncă nu există evaluări
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Infective Endocarditis Case ReportDocument40 paginiInfective Endocarditis Case Reportliu_owen17100% (1)
- Hylestad Stave ChurchDocument3 paginiHylestad Stave Churchsanjo93Încă nu există evaluări
- Parks For The People: InsideDocument5 paginiParks For The People: InsideTommy WilkinsonÎncă nu există evaluări
- Urinary System-1Document48 paginiUrinary System-1Muhammad SajjadÎncă nu există evaluări
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Shadows of Esteren The Tri-Kazelian Bestiary PDFDocument29 paginiShadows of Esteren The Tri-Kazelian Bestiary PDFAnonymous iL4zr96LKB100% (2)
- Customer Satisfaction of DairyDocument118 paginiCustomer Satisfaction of DairynitindeoraÎncă nu există evaluări
- Stages of Evolution of ManDocument3 paginiStages of Evolution of ManTristan Nialler100% (1)
- Ao 2011-0002Document7 paginiAo 2011-0002Suzette Paredes-RagosÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Contoh Soalan TOV Year 5Document8 paginiContoh Soalan TOV Year 5Fatin Nur HafizahÎncă nu există evaluări
- BG QuestsDocument71 paginiBG QuestsJeffrey DamherÎncă nu există evaluări
- Output RAISEPlanDocument4 paginiOutput RAISEPlanLovely Venia JovenÎncă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)