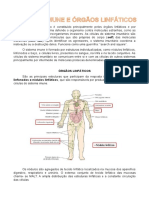
S-ar putea să vă placă și
- 08564718082016introducao A Saude Publica Aula 04Document18 pagini08564718082016introducao A Saude Publica Aula 04hosana_rrÎncă nu există evaluări
- Cárie Dentária Introdução IDocument207 paginiCárie Dentária Introdução ICibele Correia100% (1)
- Atividade 500 Pontos 29 10 2020Document9 paginiAtividade 500 Pontos 29 10 2020Luuh MacieiraÎncă nu există evaluări
- Questões para Estudo de ImunologiaDocument9 paginiQuestões para Estudo de ImunologiaauroeagÎncă nu există evaluări
- 01 - Introdução Ao Sistema ImunológicoDocument6 pagini01 - Introdução Ao Sistema ImunológicoDiego FonsecaÎncă nu există evaluări
- Estudo Dirigido de ImunologiaDocument9 paginiEstudo Dirigido de ImunologiaLuan VacarinÎncă nu există evaluări
- 10 - Tecido Imunológico e Órgãos Linfoides - MedresumosDocument4 pagini10 - Tecido Imunológico e Órgãos Linfoides - MedresumosaeadaÎncă nu există evaluări
- RPH ImunologiaDocument24 paginiRPH ImunologiarotacomandoÎncă nu există evaluări
- O Sistema ImuneDocument14 paginiO Sistema ImuneIasmyn DiasÎncă nu există evaluări
- Sistema InatoDocument10 paginiSistema InatoAlanLourençoÎncă nu există evaluări
- Sistema ImunológicoDocument34 paginiSistema ImunológiconazyalenskaÎncă nu există evaluări
- Imunopatologia Aula 2Document36 paginiImunopatologia Aula 2adilsonjacinto19945Încă nu există evaluări
- Apostila de Imunologia (Pratica)Document100 paginiApostila de Imunologia (Pratica)yurico122Încă nu există evaluări
- CHAT Sistema ImunólogicoDocument5 paginiCHAT Sistema ImunólogicoPablo PietroÎncă nu există evaluări
- Estudo Dirigido 1 GabaritoDocument3 paginiEstudo Dirigido 1 GabaritoRoseli de Santana estevamÎncă nu există evaluări
- Pós Aulas Encontro 9 Ao Encontro 15 - MicrobiologiaDocument11 paginiPós Aulas Encontro 9 Ao Encontro 15 - MicrobiologiaAssociação Jardins Roma 34 -3214-2314Încă nu există evaluări
- Resumos de ImunologiaDocument8 paginiResumos de ImunologiaJuliana LimaÎncă nu există evaluări
- Sistema Imunológico BiologiaDocument10 paginiSistema Imunológico BiologiaPedro Henrique TeixeiraÎncă nu există evaluări
- Aula 5 - Fisiologia Do Sistema Imune (Atualização)Document12 paginiAula 5 - Fisiologia Do Sistema Imune (Atualização)Lázaro Victor Santos Mendonça100% (1)
- Questoes Imunologia PDFDocument4 paginiQuestoes Imunologia PDFAna MarquesÎncă nu există evaluări
- Ed 1 Imunidade InataDocument13 paginiEd 1 Imunidade InataIsrael RodriguesÎncă nu există evaluări
- Resumo Histologia Do Sistema Imune e Órgãos LinfáticosDocument13 paginiResumo Histologia Do Sistema Imune e Órgãos LinfáticosAlexandre AugustusÎncă nu există evaluări
- Imunologia IDocument3 paginiImunologia Ifeliaraujo16Încă nu există evaluări
- Ler LerDocument106 paginiLer LerAndressa SantanaÎncă nu există evaluări
- Imunidade HumoralDocument4 paginiImunidade HumoralLaura FerreiraÎncă nu există evaluări
- ImunologiaDocument20 paginiImunologiaProf. Sândi BarrosÎncă nu există evaluări
- Imunologia Aula 1Document5 paginiImunologia Aula 1RhuanLopesÎncă nu există evaluări
- Células e Tecidos Do Sistema ImuneDocument6 paginiCélulas e Tecidos Do Sistema ImuneIsaacSousaÎncă nu există evaluări
- Sistema Imunológico - Vacinas - Ciências - Prof. JoãoDocument27 paginiSistema Imunológico - Vacinas - Ciências - Prof. JoãoEdivon OliveiraÎncă nu există evaluări
- Resumo Imunologia VeterináriaDocument103 paginiResumo Imunologia VeterináriaThaís MedeirosÎncă nu există evaluări
- Mod. II Prob. IIDocument18 paginiMod. II Prob. IIgerlan rodriguesÎncă nu există evaluări
- Cap 1 - Propriedades Gerais Das Respostas ImunesDocument5 paginiCap 1 - Propriedades Gerais Das Respostas ImunesMatheusÎncă nu există evaluări
- Resumo Imunologia VETERINÁRIADocument8 paginiResumo Imunologia VETERINÁRIAGiovanna CherubiniÎncă nu există evaluări
- Leitura II - Aula 2 - ImunologiaDocument106 paginiLeitura II - Aula 2 - ImunologiaEmanuelle ThaynáÎncă nu există evaluări
- Sistema ImunológicoDocument53 paginiSistema ImunológicoLuana DonadelÎncă nu există evaluări
- Nocoes de ImunotoxicologiaDocument22 paginiNocoes de ImunotoxicologiaSaah BatistaÎncă nu există evaluări
- 9 Ef 13 DDocument74 pagini9 Ef 13 DLaura ScottÎncă nu există evaluări
- Apostila Microbiologia e ParasitologiaDocument170 paginiApostila Microbiologia e ParasitologiaenfcarolinecavalcanteÎncă nu există evaluări
- GT2 Resolução Febre v.2.31Document18 paginiGT2 Resolução Febre v.2.31Paulo TeixeiraÎncă nu există evaluări
- Resumo Das Aulas de ImunologiaDocument56 paginiResumo Das Aulas de ImunologiaClaudio Vieira da SilvaÎncă nu există evaluări
- Imuno - Células e OrgãosDocument5 paginiImuno - Células e OrgãosKarin TreichelÎncă nu există evaluări
- ImunologiaDocument9 paginiImunologiaAlexia LimaÎncă nu există evaluări
- Sistema ImunológicoDocument32 paginiSistema ImunológicoTaciana De Oliveira Bruxel100% (5)
- ESTUDO DIRIGIDO AV2 MICRO E IMUNO 2021 2s-1 ComrespostaDocument4 paginiESTUDO DIRIGIDO AV2 MICRO E IMUNO 2021 2s-1 ComrespostaDaniel FerreiraÎncă nu există evaluări
- ImunologiaDocument10 paginiImunologiaJanine Rafael100% (1)
- (IMUNO) Imunologia Resposta Inata e Adaptativa - RESUMODocument11 pagini(IMUNO) Imunologia Resposta Inata e Adaptativa - RESUMOSamara Santos100% (1)
- Sistema ImunológicoDocument7 paginiSistema ImunológicoVisionário Wilson TavaresÎncă nu există evaluări
- Resumo 1° ProvaDocument6 paginiResumo 1° ProvaIgor KennedyÎncă nu există evaluări
- Ficha Resição Biologia Funcional - Sistema ImuneDocument18 paginiFicha Resição Biologia Funcional - Sistema ImuneDiogo Botelho Medeiros BrilhanteÎncă nu există evaluări
- Mecanismos de Agressão e Defesa PDFDocument122 paginiMecanismos de Agressão e Defesa PDFSabrina FerreiraÎncă nu există evaluări
- Medresumo 2016 - Módulo 04 - Mecanismos de Agressao e Defesa (Mad) IIDocument197 paginiMedresumo 2016 - Módulo 04 - Mecanismos de Agressao e Defesa (Mad) IIOsmair AlvesÎncă nu există evaluări
- Aula Prática - VacinaDocument4 paginiAula Prática - VacinaLeonardo LopesÎncă nu există evaluări
- Unidade I:: 1. Conceitos de ImunidadeDocument64 paginiUnidade I:: 1. Conceitos de ImunidadeGloria MariaÎncă nu există evaluări
- Sistema ImunologicoDocument5 paginiSistema Imunologicoelisangelabispo.odontoÎncă nu există evaluări
- Imunidade Específica e Não EspecíficaDocument4 paginiImunidade Específica e Não EspecíficaCarolina GonçalvesÎncă nu există evaluări
- Sistema ImunológicoDocument17 paginiSistema Imunológicojoyce santosÎncă nu există evaluări
- R - D - Ricardo Jose CaneverDocument99 paginiR - D - Ricardo Jose CaneverAnaRafaelaSilvaÎncă nu există evaluări
- Perimetro Escrotal e Idade A Puberdade em Ovinos MDocument10 paginiPerimetro Escrotal e Idade A Puberdade em Ovinos MAnaRafaelaSilvaÎncă nu există evaluări
- Melhoramento Genético Aplicado A Produção Animal - Jonas Carlos Campos Pereiro PDFDocument298 paginiMelhoramento Genético Aplicado A Produção Animal - Jonas Carlos Campos Pereiro PDFDenise GuerreiroÎncă nu există evaluări
- Abate e Processamente de Carne de AvestruzDocument56 paginiAbate e Processamente de Carne de AvestruzAnaRafaelaSilva33% (6)
- Suíno ParasitoDocument47 paginiSuíno ParasitoAnaRafaelaSilva100% (1)
- Tumor e Síndrome CarcinóideDocument49 paginiTumor e Síndrome CarcinóideSandro RolimÎncă nu există evaluări
- Plano de Ensino Biotecnologia de AlimentosDocument5 paginiPlano de Ensino Biotecnologia de AlimentossamugodÎncă nu există evaluări
- Slides - Fisiologia NeuromuscularDocument35 paginiSlides - Fisiologia NeuromuscularVitória EmanuellyÎncă nu există evaluări
- Acidentes ÓsseosDocument12 paginiAcidentes ÓsseosAlexandre Tolosa100% (2)
- A Magia Dos ChacrasDocument13 paginiA Magia Dos Chacrascatiafermon6606Încă nu există evaluări
- Farmacologia 2Document1 paginăFarmacologia 2sartsicledÎncă nu există evaluări
- Aula 1 - Aptidão Física Aplicada A Educação Física PDFDocument31 paginiAula 1 - Aptidão Física Aplicada A Educação Física PDFMarceloÎncă nu există evaluări
- Termografia BiomedicaDocument12 paginiTermografia BiomedicaTiago MarcheseÎncă nu există evaluări
- Ciências 6º - 3º CorteDocument45 paginiCiências 6º - 3º Cortesilvajeane657Încă nu există evaluări
- Direito À Vida Do NascituroDocument175 paginiDireito À Vida Do NascituroLevergerÎncă nu există evaluări
- Inadequacoes em Formulacoes de Enunciados de Atividades e Avaliacoes Escritas PDFDocument11 paginiInadequacoes em Formulacoes de Enunciados de Atividades e Avaliacoes Escritas PDFAnonymous 1FlqqXÎncă nu există evaluări
- Transicao Agroecologica Vol6 1Document358 paginiTransicao Agroecologica Vol6 1Emmanuel AlmadaÎncă nu există evaluări
- Felicita - Annalisa Volgarini PDFDocument93 paginiFelicita - Annalisa Volgarini PDFMarielle CristinaÎncă nu există evaluări
- CZZ CanoasDocument4 paginiCZZ CanoasSusy DiazÎncă nu există evaluări
- Geometria SagradaDocument116 paginiGeometria SagradaMarcela SerenoÎncă nu există evaluări
- Revista Skin Detox - Prime Pharma MagistralDocument56 paginiRevista Skin Detox - Prime Pharma MagistralTeresa Helena PiauilinoÎncă nu există evaluări
- Agenda 1 Ano Abril - MaioDocument21 paginiAgenda 1 Ano Abril - Maiorenata souzaÎncă nu există evaluări
- Simulado - Ciên - 6º AnoDocument3 paginiSimulado - Ciên - 6º AnoMATEUS FRANCISCO BERALDOÎncă nu există evaluări
- 03 e 04 - Prof Paulo Vasques - Medicina LegalDocument6 pagini03 e 04 - Prof Paulo Vasques - Medicina LegalAdriano Scott RochaÎncă nu există evaluări
- Resumo - Fisiologia Do ExercicioDocument128 paginiResumo - Fisiologia Do ExercicioMiguel AmaroÎncă nu există evaluări
- Histologia ClínicaDocument205 paginiHistologia ClínicaAdeilson ReisÎncă nu există evaluări
- Legacy of The Nine Realms 3 - Ruins of ChaosDocument645 paginiLegacy of The Nine Realms 3 - Ruins of ChaosFranciele SantosÎncă nu există evaluări
- Psicologia Do Desenvolvimento Da Criança e Do Adolescente - Unidade1Document13 paginiPsicologia Do Desenvolvimento Da Criança e Do Adolescente - Unidade1Anabela SousaÎncă nu există evaluări
- R - D - Ana Suy Sesarino Kuss PDFDocument96 paginiR - D - Ana Suy Sesarino Kuss PDFCarina Duda BittencourtÎncă nu există evaluări
- Actividade Agrcola - Abilio SousaDocument16 paginiActividade Agrcola - Abilio SousaFeliz MauricioÎncă nu există evaluări
- Fisiologia Do Exercício Físico e Hipertensão Arterial: Uma Breve IntroduçãoDocument7 paginiFisiologia Do Exercício Físico e Hipertensão Arterial: Uma Breve IntroduçãoLaila SiqueiraÎncă nu există evaluări
- Caso Clínico - CIRURGIA1Document15 paginiCaso Clínico - CIRURGIA1Gina20verÎncă nu există evaluări
- Atividade Avaliativa 6 AnoDocument5 paginiAtividade Avaliativa 6 AnoJoaoÎncă nu există evaluări
- Calor: Influencia Da Temperatura Na Saúde Do TrabalhadorDocument10 paginiCalor: Influencia Da Temperatura Na Saúde Do TrabalhadorValéria Araújo CavalcanteÎncă nu există evaluări
- MilhoDocument333 paginiMilhoEdmar TuelherÎncă nu există evaluări