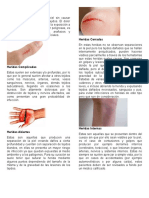
S-ar putea să vă placă și
- Sensación y percepción: umbrales, adaptación y sistemas sensorialesDocument8 paginiSensación y percepción: umbrales, adaptación y sistemas sensorialesBrailenys ReyesÎncă nu există evaluări
- Sistema Neuroendocrino y Organos de Los Sentidos PDFDocument24 paginiSistema Neuroendocrino y Organos de Los Sentidos PDFJulioCarlosMartinÎncă nu există evaluări
- Sócrates, Platón y AristótelesDocument3 paginiSócrates, Platón y Aristótelesjuan muñozÎncă nu există evaluări
- Colocacion FamiliarDocument3 paginiColocacion FamiliarmaurobarretoÎncă nu există evaluări
- La CleptomaniaDocument6 paginiLa CleptomaniaSylvia NuñezÎncă nu există evaluări
- CCT Gobernacion Guarico 2014-2016 PDFDocument33 paginiCCT Gobernacion Guarico 2014-2016 PDFKeith LedezmaÎncă nu există evaluări
- ALCANCE Y JUSTIFICACIÓN DE LA MEDICINA LEGAL EN VENEZUELADocument20 paginiALCANCE Y JUSTIFICACIÓN DE LA MEDICINA LEGAL EN VENEZUELAyenny salcedoÎncă nu există evaluări
- Ensayo Derecho AgrarioDocument9 paginiEnsayo Derecho AgrariojecciÎncă nu există evaluări
- Clase de Embriología Del SNCDocument19 paginiClase de Embriología Del SNCLuis Angel Unocc GuillenÎncă nu există evaluări
- Etica y Gobierno, Valores y Principios de La CRBVDocument17 paginiEtica y Gobierno, Valores y Principios de La CRBVJoseRojas100% (2)
- Atencion A La VictimaDocument6 paginiAtencion A La VictimaAshley RiveroÎncă nu există evaluări
- Pares Craneales..Document20 paginiPares Craneales..Paola FigueroaÎncă nu există evaluări
- Funciones psíquicas y sus alteracionesDocument4 paginiFunciones psíquicas y sus alteracionesOlga ParchutikÎncă nu există evaluări
- Trabajo Pares CranealesDocument54 paginiTrabajo Pares CranealesLuulaa Minguin GarciaÎncă nu există evaluări
- Contrato CanaimaDocument2 paginiContrato CanaimaJAIRO PILDAINÎncă nu există evaluări
- Desarrollo fetal y gestaciónDocument4 paginiDesarrollo fetal y gestaciónMichael Andrés IndÎncă nu există evaluări
- Vasculitis Infecciosa y No InfecciosaDocument33 paginiVasculitis Infecciosa y No InfecciosaDaniellis Choles QuinteroÎncă nu există evaluări
- Importancia del método clínico en la medicinaDocument16 paginiImportancia del método clínico en la medicinaAnonymous WTGcrVÎncă nu există evaluări
- Fiebre Purpurica BrasileñaDocument3 paginiFiebre Purpurica BrasileñaRossi50% (2)
- MPPSPDocument65 paginiMPPSPRiger GomezÎncă nu există evaluări
- Los Venezolanos Están DesculturizadosDocument7 paginiLos Venezolanos Están DesculturizadosNilia RamirezÎncă nu există evaluări
- Historia PatologiaDocument4 paginiHistoria PatologiaJose SirittÎncă nu există evaluări
- 4643 D Libro AdopcionDocument201 pagini4643 D Libro AdopcionAbel A Ochoa SuárezÎncă nu există evaluări
- Educar en y para Los Derechos Humanos AnneliseDocument6 paginiEducar en y para Los Derechos Humanos AnneliseMELIZ1981Încă nu există evaluări
- Trabajo de CognicionDocument20 paginiTrabajo de CognicionSofi LopezÎncă nu există evaluări
- Ensayo HCDocument2 paginiEnsayo HCDANIELA LUNA RAMOSÎncă nu există evaluări
- 3ra Actividad Investigacion. MEDICINA LEGALDocument20 pagini3ra Actividad Investigacion. MEDICINA LEGALAnonymous pLcJP2jbÎncă nu există evaluări
- Infome Medico Legal 1Document6 paginiInfome Medico Legal 1JaimeÎncă nu există evaluări
- Trata y Tráfico de Personas (Conceptos, Normativa y Situación Actual)Document16 paginiTrata y Tráfico de Personas (Conceptos, Normativa y Situación Actual)Valentin GarciaÎncă nu există evaluări
- Historia de La DactilosDocument63 paginiHistoria de La DactilosÓscar Javier Bustos LeónÎncă nu există evaluări
- Tijera de MayoDocument12 paginiTijera de MayoAccessobethelLACelulaÎncă nu există evaluări
- Desarrollo Del Potencial HumanoDocument4 paginiDesarrollo Del Potencial Humanojerigpargas100% (1)
- Historia de La Medicina LegalDocument27 paginiHistoria de La Medicina Legaljose gonzalez0% (1)
- Infografia MiguelDocument4 paginiInfografia Miguelcentevi zalezgonÎncă nu există evaluări
- CONANI Revista 1Document40 paginiCONANI Revista 1CONANIRDoÎncă nu există evaluări
- Integración de Las Vías Nerviosas GeneralesDocument40 paginiIntegración de Las Vías Nerviosas GeneralesJavier CastroÎncă nu există evaluări
- 10 Formato Control Cumplimiento Guardia Pnfa PF SenamecfDocument2 pagini10 Formato Control Cumplimiento Guardia Pnfa PF SenamecfnoblezadeolmosÎncă nu există evaluări
- Desarrollo Personal Corrientes y EnfoquesDocument3 paginiDesarrollo Personal Corrientes y EnfoquesYanira Marielena Alvarez CruzÎncă nu există evaluări
- Generalidades Del Sistema NerviosoDocument97 paginiGeneralidades Del Sistema NerviosoRafael AlvarezÎncă nu există evaluări
- Sustancias facilitadoras de asalto sexual en BogotáDocument116 paginiSustancias facilitadoras de asalto sexual en BogotáJose David LopezÎncă nu există evaluări
- Ensayo El Cerebro y La CogniciónDocument4 paginiEnsayo El Cerebro y La CogniciónverlyfrancdÎncă nu există evaluări
- Paper Tomás de Aquino EpistemologiaDocument5 paginiPaper Tomás de Aquino Epistemologiaekimmoreno100% (2)
- Alternativas A La Prosecucion Del ProcesoDocument12 paginiAlternativas A La Prosecucion Del ProcesoNataly VelasquezÎncă nu există evaluări
- Ensayo Hemisferios DOCTORADO OSCAR MAMANI QUISPEDocument4 paginiEnsayo Hemisferios DOCTORADO OSCAR MAMANI QUISPEOscarÎncă nu există evaluări
- Comportamiento o Humano en La Sociedad, Diferencia Del Comportamiento Individual Colectivo y SociedadDocument3 paginiComportamiento o Humano en La Sociedad, Diferencia Del Comportamiento Individual Colectivo y SociedadMacz Eugenio YatÎncă nu există evaluări
- ArticulacionDocument3 paginiArticulacionhenrryÎncă nu există evaluări
- Trabajo CriminalísticaDocument10 paginiTrabajo CriminalísticaPaula Andrea Torres Camacho100% (1)
- Ensayo de CriminologiaDocument4 paginiEnsayo de CriminologiaEsther SanchezÎncă nu există evaluări
- Sensacion y PercepcionDocument3 paginiSensacion y PercepcioneliudÎncă nu există evaluări
- Desarrollo Prenatal y GeneticaDocument19 paginiDesarrollo Prenatal y GeneticaJennifer Molano Diaz100% (1)
- Practica Agudeza Visual y AuditivaDocument15 paginiPractica Agudeza Visual y AuditivaLaura JimenezÎncă nu există evaluări
- Medicina Legal - Infanticidio, Suicidio y HomicidioDocument20 paginiMedicina Legal - Infanticidio, Suicidio y HomicidioRoger Alejandro MontoyaÎncă nu există evaluări
- Tesis Kimberlin y DeibyDocument39 paginiTesis Kimberlin y DeibyYurbeilyn100% (1)
- Genetica Forense y Problemas de Paternidad DiscutidaDocument18 paginiGenetica Forense y Problemas de Paternidad DiscutidaEvelyn Calderón Martínez100% (5)
- MEDULA ESPINAL-MONOGRAFÍA FinalDocument15 paginiMEDULA ESPINAL-MONOGRAFÍA FinalIvan PumaÎncă nu există evaluări
- D1EADocument1 paginăD1EAJesús Alejandro RondónÎncă nu există evaluări
- Inspección balística femeninaDocument9 paginiInspección balística femeninaLisandroÎncă nu există evaluări
- Conducta y Corteza Cerebral Sistema Limbico y HormonasDocument5 paginiConducta y Corteza Cerebral Sistema Limbico y HormonasFernanda Reyes26Încă nu există evaluări
- Bases Neurofisiologicas Del AprendizajeDocument12 paginiBases Neurofisiologicas Del AprendizajeEliana ConchaÎncă nu există evaluări
- Organizacion Funcional de La Corteza Cerebral - Dr. Brusco D. Marquez CLASE 2 PDFDocument3 paginiOrganizacion Funcional de La Corteza Cerebral - Dr. Brusco D. Marquez CLASE 2 PDFEduardo Fasce100% (1)
- NeuroDocument13 paginiNeuroUniversidad Cooperativa Sede Neiva/HuilaÎncă nu există evaluări
- Proyecto de Aula. Giovanna, Anyela, Juan David, ChristianDocument9 paginiProyecto de Aula. Giovanna, Anyela, Juan David, ChristianUniversidad Cooperativa Sede Neiva/HuilaÎncă nu există evaluări
- Proyecto de AulaDocument3 paginiProyecto de AulaUniversidad Cooperativa Sede Neiva/HuilaÎncă nu există evaluări
- Blog NeurocienciasDocument26 paginiBlog NeurocienciasUniversidad Cooperativa Sede Neiva/HuilaÎncă nu există evaluări
- GFHGDHYDocument6 paginiGFHGDHYyessenia oscoÎncă nu există evaluări
- Tipos de VolcanDocument3 paginiTipos de VolcanMcarmen AlemanyÎncă nu există evaluări
- Geología del Perú: Evolución geológica y yacimientos mineralesDocument157 paginiGeología del Perú: Evolución geológica y yacimientos mineralesWalter Orlando Boñón AlcántaraÎncă nu există evaluări
- La Radiestesia y La Red HartmannDocument6 paginiLa Radiestesia y La Red HartmannJose Marin100% (1)
- Adaptacion CCSS 1 EsoDocument3 paginiAdaptacion CCSS 1 EsomariviprofeÎncă nu există evaluări
- Cuadro Comparativo Vigostky Piaget WallonDocument6 paginiCuadro Comparativo Vigostky Piaget WallonLucila Rios DiazÎncă nu există evaluări
- Aislamiento de microorganismos en gimnasio universidadDocument6 paginiAislamiento de microorganismos en gimnasio universidadMaría José Díaz RuizÎncă nu există evaluări
- Derecho y Conflicto EntelmanDocument90 paginiDerecho y Conflicto EntelmanLeanaKowalczukÎncă nu există evaluări
- Prueba Unidad 1 Estructura y Funcionamiento de Los Seres VivosDocument3 paginiPrueba Unidad 1 Estructura y Funcionamiento de Los Seres Vivoskarinapatri25hotmailcomÎncă nu există evaluări
- Manual Arcgis 10.2Document6 paginiManual Arcgis 10.2Jean Querevalú LazaroÎncă nu există evaluări
- cABO ORTEGALDocument3 paginicABO ORTEGALCésar Antonio Pardo RipollÎncă nu există evaluări
- A Cidade Dos BarriosDocument39 paginiA Cidade Dos BarriosJuan CarlosÎncă nu există evaluări
- Monografía Corte y Relleno PDFDocument68 paginiMonografía Corte y Relleno PDFEdwin Joel Quispe100% (9)
- FotosintesisDocument1 paginăFotosintesiszero1202Încă nu există evaluări
- Depositos Tipo CARLINDocument12 paginiDepositos Tipo CARLINABRAHAMÎncă nu există evaluări
- Sistema Muscular 2016Document38 paginiSistema Muscular 2016Cesar AlejandroÎncă nu există evaluări
- ChorreraDocument10 paginiChorreraAlexandraMarroneÎncă nu există evaluări
- Fluidos MineralizantesDocument6 paginiFluidos MineralizantesDenis Avendaño NinaÎncă nu există evaluări
- Batolito de Patáz: Características geológicas y mineralizaciónDocument26 paginiBatolito de Patáz: Características geológicas y mineralizaciónErika Cruzado Merino100% (1)
- Manual DuraznoDocument42 paginiManual DuraznoRocio Morales Vargas100% (2)
- Clasificación de la porosidad en rocas carbonatadas según LuciaDocument5 paginiClasificación de la porosidad en rocas carbonatadas según LuciaSamuel P. García100% (1)
- Parcelas ColbergDocument85 paginiParcelas Colbergrikifreytes100% (2)
- Corrosión Experimento de EvansDocument6 paginiCorrosión Experimento de EvansJairo AlvarezÎncă nu există evaluări
- WWC Formulas U CapacidadesDocument48 paginiWWC Formulas U CapacidadesGustavo Gomez MolinaÎncă nu există evaluări
- Colada de LavaDocument3 paginiColada de LavaIng EstebanÎncă nu există evaluări
- Proyecto de Pepino Dr. OTTONIELDocument27 paginiProyecto de Pepino Dr. OTTONIELDesiree BetancourtÎncă nu există evaluări
- Metodos de Separacion y Purificacion de Compuestos Organicos Por CromatografiaDocument4 paginiMetodos de Separacion y Purificacion de Compuestos Organicos Por CromatografiafranciscoÎncă nu există evaluări
- Producción avícola: subsistemas y procesos claveDocument20 paginiProducción avícola: subsistemas y procesos clavestefanyÎncă nu există evaluări
- Biología y geología: La célula, unidad de vidaDocument27 paginiBiología y geología: La célula, unidad de vidalourdesÎncă nu există evaluări
- Nectar de DuraznoDocument21 paginiNectar de DuraznojairoÎncă nu există evaluări