S-ar putea să vă placă și
- cells: Role of TGF-β in Skin Chronic Wounds: A Keratinocyte PerspectiveDocument17 paginicells: Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspectiveoni handayaniÎncă nu există evaluări
- 1 s2.0 S0014579305006319 MainDocument8 pagini1 s2.0 S0014579305006319 MainKarthik AripakaÎncă nu există evaluări
- 1995 Arvanitakis JimmunolDocument10 pagini1995 Arvanitakis JimmunolleandrosarvanitakisÎncă nu există evaluări
- Transforming Growth Factor-In Development and Differentiation of Stem CellsDocument17 paginiTransforming Growth Factor-In Development and Differentiation of Stem CellsJAN CAMILLE OLIVARESÎncă nu există evaluări
- Slavin 1996Document6 paginiSlavin 1996jahdsdjad asffdhsajhajdkÎncă nu există evaluări
- 2) Targeting the TGFβ signalling ReviewDocument23 pagini2) Targeting the TGFβ signalling Reviewkani55Încă nu există evaluări
- The Role of TGFB Signaling in Wound Epithelialization: Horacio Ramirez, Shailee B. Patel, and Irena PastarDocument10 paginiThe Role of TGFB Signaling in Wound Epithelialization: Horacio Ramirez, Shailee B. Patel, and Irena PastarYodha PranataÎncă nu există evaluări
- Transglutaminase 2 in The Balance of Cell Death and SurvivalDocument6 paginiTransglutaminase 2 in The Balance of Cell Death and SurvivalOsman FrankÎncă nu există evaluări
- Topic:: TGF Beta Signaling PathwayDocument15 paginiTopic:: TGF Beta Signaling PathwayMehwish AhmedÎncă nu există evaluări
- TGF-β Mediated SMAD Signaling NotesDocument5 paginiTGF-β Mediated SMAD Signaling Noteskim.yeuÎncă nu există evaluări
- TGF-beta Rev 2019Document34 paginiTGF-beta Rev 2019Banchob SripaÎncă nu există evaluări
- Upregulation of the Wnt/ β-catenin Pathway Induced by Transforming Growth Factor-β in Hypertrophic Scars and KeloidsDocument8 paginiUpregulation of the Wnt/ β-catenin Pathway Induced by Transforming Growth Factor-β in Hypertrophic Scars and KeloidsNayapuramesh BabuÎncă nu există evaluări
- Best of TGB and ImunologyDocument22 paginiBest of TGB and ImunologyApriyanto LatiefÎncă nu există evaluări
- Targeting TGF β signal transduction for cancer therapyDocument20 paginiTargeting TGF β signal transduction for cancer therapyIgnacio martinÎncă nu există evaluări
- TGF Beta Signaling Pathway: MechanismDocument7 paginiTGF Beta Signaling Pathway: MechanismAkbaruddin SholehÎncă nu există evaluări
- Artigo OncoDocument14 paginiArtigo OncoCatarina LuísÎncă nu există evaluări
- New Insights Into The Functions of Intersectin-1s: Sanda Predescu, Cristina Bardita, and Dan PredescuDocument3 paginiNew Insights Into The Functions of Intersectin-1s: Sanda Predescu, Cristina Bardita, and Dan PredescuSergeat18BÎncă nu există evaluări
- PIIS0960982208008221Document9 paginiPIIS0960982208008221melcapellaÎncă nu există evaluări
- Tgf-Health and Disease: Action in The Cartilage inDocument21 paginiTgf-Health and Disease: Action in The Cartilage inResa PutraÎncă nu există evaluări
- EGF TGF & B ReceptorsDocument22 paginiEGF TGF & B ReceptorsFatima IffatÎncă nu există evaluări
- An Integrative View On The Role of TGF-B in TheDocument6 paginiAn Integrative View On The Role of TGF-B in Theagustinaw1981Încă nu există evaluări
- Mini EW: Department Laboratory Medicine and Pathology, Muyo Graduate School Medicine, Rochester, MinnesotaDocument14 paginiMini EW: Department Laboratory Medicine and Pathology, Muyo Graduate School Medicine, Rochester, MinnesotaMARIELLA AMANDA RAMOS VALDIVIEZOÎncă nu există evaluări
- GADD45B Growth Arrest and DNA-damage-inducible, BetaDocument6 paginiGADD45B Growth Arrest and DNA-damage-inducible, BetaHassan Ahmed HassanÎncă nu există evaluări
- Galunisertib DetailsDocument19 paginiGalunisertib DetailsRahul DasÎncă nu există evaluări
- Field 等 - 2020 - Mitochondrial Integrity Regulated by Lipid MetabolDocument32 paginiField 等 - 2020 - Mitochondrial Integrity Regulated by Lipid MetabolliqinggreenÎncă nu există evaluări
- ENDOSIGN Review v0 EdmvDocument33 paginiENDOSIGN Review v0 EdmvMavi Camarasa SabaterÎncă nu există evaluări
- Trevor Nong Final ProjectDocument7 paginiTrevor Nong Final Projectapi-375199340Încă nu există evaluări
- mTOR Signaling Interactive PathwayDocument6 paginimTOR Signaling Interactive PathwayKiki GirindraÎncă nu există evaluări
- Alfa TomatinaDocument11 paginiAlfa TomatinaSava1988Încă nu există evaluări
- CKD - Signaling in fibrosis TGF-β, WNT, and YAP-TAZ convergeDocument14 paginiCKD - Signaling in fibrosis TGF-β, WNT, and YAP-TAZ convergeCher IshÎncă nu există evaluări
- Articulo de Investigacion 1Document20 paginiArticulo de Investigacion 1HÉCTOR MEDINAÎncă nu există evaluări
- For Translation - Fr.enDocument11 paginiFor Translation - Fr.enBoutaina AddoumÎncă nu există evaluări
- TGF - Signaling in The Kidney: Profibrotic and Protective EffectsDocument11 paginiTGF - Signaling in The Kidney: Profibrotic and Protective EffectsMariel AbatayoÎncă nu există evaluări
- tmp2F1F TMPDocument16 paginitmp2F1F TMPFrontiersÎncă nu există evaluări
- TGF-B Signaling Pathway Inactivation and Cell Cycle Deregulation in The Development of Gastric Cancer Role of The Spectrin ELFDocument8 paginiTGF-B Signaling Pathway Inactivation and Cell Cycle Deregulation in The Development of Gastric Cancer Role of The Spectrin ELFVetPhoenix StkÎncă nu există evaluări
- Growth Factor Receptor Bound Protein2Document2 paginiGrowth Factor Receptor Bound Protein2rajeshmangalÎncă nu există evaluări
- Smad4 Loss Enables Egf, Tgfβ1 And S100A8/A9 Induced Activation Of Critical Pathways To Invasion In Human Pancreatic Adenocarcinoma CellsDocument18 paginiSmad4 Loss Enables Egf, Tgfβ1 And S100A8/A9 Induced Activation Of Critical Pathways To Invasion In Human Pancreatic Adenocarcinoma CellsSisca Dwi AgustinaÎncă nu există evaluări
- Application of Recombinant DNATechnologies On Sub-Cloning of Transcriptional Co-FactorDocument6 paginiApplication of Recombinant DNATechnologies On Sub-Cloning of Transcriptional Co-FactorInternational Journal of Innovative Science and Research TechnologyÎncă nu există evaluări
- Anti-Tumoral Effects of Mir-3189-3P in GlioblastomaDocument14 paginiAnti-Tumoral Effects of Mir-3189-3P in GlioblastomaAdlerÎncă nu există evaluări
- Hippo CancerDocument7 paginiHippo CancerMaria Elena MaldonadoÎncă nu există evaluări
- Delayed Wound Healing in Immunode®cient TGF-b1 Knockout MiceDocument9 paginiDelayed Wound Healing in Immunode®cient TGF-b1 Knockout MiceMarcelo EsquisattoÎncă nu există evaluări
- Metformin inhibits TGF-β1-induced epithelial-to-mesenchymal transition like processDocument13 paginiMetformin inhibits TGF-β1-induced epithelial-to-mesenchymal transition like processkartikawensdiÎncă nu există evaluări
- Thyroid CA Therapeutic TargetDocument12 paginiThyroid CA Therapeutic TargetJoni MokodoÎncă nu există evaluări
- Zanottofilho 2010Document12 paginiZanottofilho 2010Matheus Becker FreitasÎncă nu există evaluări
- Pathogenesis of Veterinary DiseaseDocument21 paginiPathogenesis of Veterinary DiseaseRandy Butternubs67% (6)
- Jak Stat InhibitorsDocument64 paginiJak Stat InhibitorsSupratim KarmakarÎncă nu există evaluări
- Myocardin Enhances Smad3-Mediated Transforming Growth Factor-β1 Signaling in a CArG Box-Independent MannerDocument9 paginiMyocardin Enhances Smad3-Mediated Transforming Growth Factor-β1 Signaling in a CArG Box-Independent ManneraneerajapÎncă nu există evaluări
- Bagus BangetDocument12 paginiBagus BangetSyiefa RenandaÎncă nu există evaluări
- Kidney InternationalDocument2 paginiKidney InternationalTweenkyy DiÎncă nu există evaluări
- Growth Factors and Growth Factor Receptors in Cancer: T. RajkumarDocument7 paginiGrowth Factors and Growth Factor Receptors in Cancer: T. RajkumarMohammed AladhraeiÎncă nu există evaluări
- BSBT612 19.03.2020 Unit 5Document5 paginiBSBT612 19.03.2020 Unit 5Siddhant SoniÎncă nu există evaluări
- Thomas 2015Document7 paginiThomas 2015Mehedi HossainÎncă nu există evaluări
- MicrobiologieDocument13 paginiMicrobiologieWafaa ZagharyÎncă nu există evaluări
- The Role of Matrix Stiffness in Hepatic Stellate.19Document4 paginiThe Role of Matrix Stiffness in Hepatic Stellate.19Piyush DagaÎncă nu există evaluări
- CKD - WNT and TGFb1 PathwaysDocument26 paginiCKD - WNT and TGFb1 PathwaysCher IshÎncă nu există evaluări
- Clinical Signi Cance of Oncogenic KIT and PDGFRA Mutations in Gastrointestinal Stromal Tumours PDFDocument22 paginiClinical Signi Cance of Oncogenic KIT and PDGFRA Mutations in Gastrointestinal Stromal Tumours PDFCarla CarolineÎncă nu există evaluări
- 2010 Articulo TGF-b1 and ROS Tobar Mol Cell Biochem 2010-LibreDocument8 pagini2010 Articulo TGF-b1 and ROS Tobar Mol Cell Biochem 2010-LibretirtasetiawanÎncă nu există evaluări
- PI3K/AKT/mTOR Pathway in Angiogenesis: Molecular NeuroscienceDocument8 paginiPI3K/AKT/mTOR Pathway in Angiogenesis: Molecular NeuroscienceAbu KhairÎncă nu există evaluări
- Jurnal2 PDFDocument12 paginiJurnal2 PDFRINDA HERNISÎncă nu există evaluări
- Step by Step GuidehxtfhDocument6 paginiStep by Step GuidehxtfhArjun NarangÎncă nu există evaluări
- Failure FPDDocument7 paginiFailure FPDArjun NarangÎncă nu există evaluări
- Retraction TechniquesDocument4 paginiRetraction TechniquesArjun NarangÎncă nu există evaluări
- Journal Pre-ProofDocument17 paginiJournal Pre-ProofArjun NarangÎncă nu există evaluări
- Removable Prosthodontics: Louis Blatterfein S. Howard PayneDocument6 paginiRemovable Prosthodontics: Louis Blatterfein S. Howard PayneArjun NarangÎncă nu există evaluări
- Comparative Prospective Study On Splint Therapy of Anterior Disc Displacement Without ReductionDocument6 paginiComparative Prospective Study On Splint Therapy of Anterior Disc Displacement Without ReductionArjun NarangÎncă nu există evaluări
- Is There A Benefit of Using An Arbitrary Facebow For The Fabrication of A Stabilization Appliance?Document7 paginiIs There A Benefit of Using An Arbitrary Facebow For The Fabrication of A Stabilization Appliance?Arjun NarangÎncă nu există evaluări
- Book Reviews: 3D Head and Neck Anatomy For Dentistry (DVD)Document2 paginiBook Reviews: 3D Head and Neck Anatomy For Dentistry (DVD)Arjun NarangÎncă nu există evaluări
- Nohre-Solvang CarlssonJPPA2015Document8 paginiNohre-Solvang CarlssonJPPA2015Arjun NarangÎncă nu există evaluări
- Intellidrug Implant For Medicine Delivery in Alzheimer'S Disease TreatmentDocument5 paginiIntellidrug Implant For Medicine Delivery in Alzheimer'S Disease TreatmentArjun NarangÎncă nu există evaluări
- Ca LeveDocument7 paginiCa LeveArjun NarangÎncă nu există evaluări
- Nutrition and Oral Health PDFDocument12 paginiNutrition and Oral Health PDFArjun NarangÎncă nu există evaluări
- Research: Journal Ov Dentistry, 8:709-720, 1958Document15 paginiResearch: Journal Ov Dentistry, 8:709-720, 1958Arjun NarangÎncă nu există evaluări
- Elastic Impression Materials:: HydrocolloidsDocument36 paginiElastic Impression Materials:: HydrocolloidsArjun NarangÎncă nu există evaluări
- 2010 AnsDocument37 pagini2010 AnsArjun NarangÎncă nu există evaluări
- 22march Uppg Questions Recalls and Answers: Connect RXPG With Twitter and Get Instant Bonus Credits! RXPG Uppg ForumDocument17 pagini22march Uppg Questions Recalls and Answers: Connect RXPG With Twitter and Get Instant Bonus Credits! RXPG Uppg ForumArjun NarangÎncă nu există evaluări
- TMJ PiyaDocument6 paginiTMJ PiyaArjun Narang100% (1)
- Prostho SR 2015Document34 paginiProstho SR 2015Arjun NarangÎncă nu există evaluări
- TestDocument14 paginiTestArjun NarangÎncă nu există evaluări
- Endo SR 2013 PDFDocument19 paginiEndo SR 2013 PDFArjun NarangÎncă nu există evaluări
- Skin - Structure and Function Flashcards - QuizletDocument5 paginiSkin - Structure and Function Flashcards - QuizletDani AnyikaÎncă nu există evaluări
- Microbiota Therapy Acts via a Regulatory T Cell MyD88:RORγt Pathway to Suppress Food AllergyDocument29 paginiMicrobiota Therapy Acts via a Regulatory T Cell MyD88:RORγt Pathway to Suppress Food AllergyFelipe Gálvez JirónÎncă nu există evaluări
- Drug Study NubainDocument2 paginiDrug Study NubainampalÎncă nu există evaluări
- Summer Training For Speed: Alwyn Cosgrove's Training Design ProgramDocument6 paginiSummer Training For Speed: Alwyn Cosgrove's Training Design ProgramSteve HeywoodÎncă nu există evaluări
- A Study On Employee MotivationDocument113 paginiA Study On Employee Motivationk eswari100% (2)
- Imbalanced Nutrition Less Than Body Requirements Diabetes NCP PDFDocument5 paginiImbalanced Nutrition Less Than Body Requirements Diabetes NCP PDFashamy acolÎncă nu există evaluări
- MRCS Revision Guide Trunk and Thorax by Mazyar Kanani Leanne Harling (Z-LiDocument184 paginiMRCS Revision Guide Trunk and Thorax by Mazyar Kanani Leanne Harling (Z-LiRahmaÎncă nu există evaluări
- Penyakit Jantung KongenitalDocument30 paginiPenyakit Jantung KongenitalrintiikÎncă nu există evaluări
- Hemolytic Uremic SyndromeDocument1 paginăHemolytic Uremic SyndromeAndrea TeranÎncă nu există evaluări
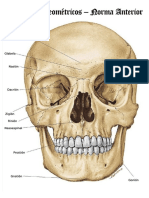
- Puntos Craneometricos CompressDocument13 paginiPuntos Craneometricos CompressLaura PÎncă nu există evaluări
- OB Case StudyDocument18 paginiOB Case StudyPramod A.NÎncă nu există evaluări
- HSCRP Level-0,5Document10 paginiHSCRP Level-0,5Nova SipahutarÎncă nu există evaluări
- Lecture 11. Avian Nervous SystemDocument6 paginiLecture 11. Avian Nervous SystemCube MaximÎncă nu există evaluări
- Anatomy OutlineDocument6 paginiAnatomy OutlineLena KhayoÎncă nu există evaluări
- 07 Bibliography FormatDocument14 pagini07 Bibliography FormatArjun KafleÎncă nu există evaluări
- Bio Lab ReportDocument7 paginiBio Lab Reportjoylee05Încă nu există evaluări
- Lecture 1 Anatomy of Prokarotic Genome 31 Jan 2012Document29 paginiLecture 1 Anatomy of Prokarotic Genome 31 Jan 2012Paramvir MannÎncă nu există evaluări
- Isometric Power Revolution Part2Document86 paginiIsometric Power Revolution Part2Smriti Ichopra100% (15)
- Worksheet 2 - Hematology - 2020 (11th Ed)Document3 paginiWorksheet 2 - Hematology - 2020 (11th Ed)ahmadmahdi18mÎncă nu există evaluări
- (Criminalistics 5) Police Polygraphy (Lie-Detection)Document15 pagini(Criminalistics 5) Police Polygraphy (Lie-Detection)Yam P Bustamante100% (2)
- Bowel Nosodes Power PointDocument15 paginiBowel Nosodes Power PointTheresa Dale83% (6)
- Chapter 18 - Apoptosis - 112612Document28 paginiChapter 18 - Apoptosis - 112612Anonymous nkR1PhIHÎncă nu există evaluări
- Resp AssessmentDocument19 paginiResp AssessmentNURSES- NOOK & CORNERÎncă nu există evaluări
- As Biology With Stafford Unit 3 Workbook Answers PDFDocument21 paginiAs Biology With Stafford Unit 3 Workbook Answers PDFFatma ZorluÎncă nu există evaluări
- Occult Meaning of Colours Purple Haze, All in My BrainDocument54 paginiOccult Meaning of Colours Purple Haze, All in My BrainrobinhoodlumÎncă nu există evaluări
- Operating RoomDocument81 paginiOperating Roomjaypee01100% (3)
- PP03L039 - Disorders of The PancreasDocument41 paginiPP03L039 - Disorders of The Pancreasapi-3805855100% (3)
- Bloody Tears Following Lefort 1Document3 paginiBloody Tears Following Lefort 1paulwhite1234Încă nu există evaluări
- Drug StudyDocument3 paginiDrug StudyStephannie MirandaÎncă nu există evaluări
- Oriflamebeautyacademy SkincareDocument173 paginiOriflamebeautyacademy Skincareapi-37629474750% (2)