S-ar putea să vă placă și
- Seminario de Investigación Estrés y CautiverioDocument26 paginiSeminario de Investigación Estrés y CautiverioAndrés L. Müller GonzálezÎncă nu există evaluări
- Joortizv,+09 Art05Document6 paginiJoortizv,+09 Art05imaÎncă nu există evaluări
- Estres e Inmunologia - EscritoDocument5 paginiEstres e Inmunologia - Escritocarlos pradaÎncă nu există evaluări
- Cronobiología: Correlatos Básicos y Médicos: Chronobiology: The Basics and Medical CorrelatesDocument7 paginiCronobiología: Correlatos Básicos y Médicos: Chronobiology: The Basics and Medical Correlatesmanu98kkskskskskiwyhÎncă nu există evaluări
- La Neuroquímica Del Estrés y El Papel de Los OpioidesDocument8 paginiLa Neuroquímica Del Estrés y El Papel de Los Opioides-Vladimir Allex-Încă nu există evaluări
- Neuroquímica Del Estrés (SECURED) PDFDocument9 paginiNeuroquímica Del Estrés (SECURED) PDFPaola RangelÎncă nu există evaluări
- Cronobiologia FISIOLOGIADocument7 paginiCronobiologia FISIOLOGIAPauleth EspinozaÎncă nu există evaluări
- Cronobiología Correlatos Basicos y MedicosDocument7 paginiCronobiología Correlatos Basicos y MedicosMa Esther R. PonceÎncă nu există evaluări
- Estres, Bienestar y Productividad en Ganado BovinoDocument5 paginiEstres, Bienestar y Productividad en Ganado BovinoGema100% (1)
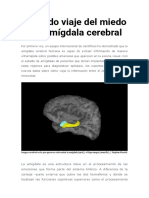
- El Rápido Viaje Del Miedo en La Amígdala CerebralDocument11 paginiEl Rápido Viaje Del Miedo en La Amígdala CerebralRomina BaezaÎncă nu există evaluări
- BASE DEL TRABAJO SELECCIONADODocument18 paginiBASE DEL TRABAJO SELECCIONADONelly ChourioÎncă nu există evaluări
- Neuroinmunoendocrinologia PDFDocument14 paginiNeuroinmunoendocrinologia PDFJulian100% (1)
- Estrés en PsiconeuroendocrinoinmunologíaDocument7 paginiEstrés en PsiconeuroendocrinoinmunologíaJorge Guerrero RodríguezÎncă nu există evaluări
- Textos Fisiologia TraducidoDocument29 paginiTextos Fisiologia Traducidoandoni_losada3501Încă nu există evaluări
- Estrés y eje hipotalámicoDocument4 paginiEstrés y eje hipotalámicoVictor RubioÎncă nu există evaluări
- Neurobiología Del Comportamiento Animal - BM EditoresDocument13 paginiNeurobiología Del Comportamiento Animal - BM EditoresCatalina Díaz MonsalveÎncă nu există evaluări
- Respuesta Metabólica Al Estrés en Pacientes Adultos Con TraumasDocument3 paginiRespuesta Metabólica Al Estrés en Pacientes Adultos Con TraumasEl Ser de LuzÎncă nu există evaluări
- Actualización: Mecanismos Celulares en Respuesta Al Estrés: SirtuinasDocument12 paginiActualización: Mecanismos Celulares en Respuesta Al Estrés: SirtuinasMonroy FernandoÎncă nu există evaluări
- Fisiopatologia Informe de Estres y AdaptacionDocument8 paginiFisiopatologia Informe de Estres y Adaptacionrafael telezÎncă nu există evaluări
- Neurobiología del estrés+Document12 paginiNeurobiología del estrés+Manel EMÎncă nu există evaluări
- 4 - EstrésDocument3 pagini4 - EstrésJJA VÎncă nu există evaluări
- Respuesta Al Estrés Actualidades en El Eje Psico-neuro-Inmunoendocrino en Enfermedades Agudas y CrónicasDocument10 paginiRespuesta Al Estrés Actualidades en El Eje Psico-neuro-Inmunoendocrino en Enfermedades Agudas y Crónicasjose venaÎncă nu există evaluări
- Grupo4 FisioanimalDocument8 paginiGrupo4 FisioanimalCarmita Noemi CastroÎncă nu există evaluări
- Estrés y Periodontitis PDFDocument10 paginiEstrés y Periodontitis PDFAngel Raúl CentenoÎncă nu există evaluări
- Carga Alostatica Parte 1Document72 paginiCarga Alostatica Parte 1AMCArroyo100% (1)
- Eje Hipotálamo-Hipofisario-AdrenalDocument5 paginiEje Hipotálamo-Hipofisario-AdrenalSilvia Valle lomas100% (1)
- Audisio E. (2020) - Interacciones SN, SE y SI (Ficha de Cátedra)Document9 paginiAudisio E. (2020) - Interacciones SN, SE y SI (Ficha de Cátedra)Augusto VillarÎncă nu există evaluări
- Homeostasis BioquimicaDocument7 paginiHomeostasis BioquimicaBernardo MoralesÎncă nu există evaluări
- Tarea 4 - Ecofisiología y Su Importancia en Producción Animal 1Document12 paginiTarea 4 - Ecofisiología y Su Importancia en Producción Animal 1Haiberth DoradoÎncă nu există evaluări
- PP XVII - Vol III Nuestro OrganismoDocument36 paginiPP XVII - Vol III Nuestro OrganismocirculodesanadorasÎncă nu există evaluări
- EcofisiologiaDocument8 paginiEcofisiologiaAngie DiazÎncă nu există evaluări
- Respuesta Metabólica Al TraumaDocument62 paginiRespuesta Metabólica Al TraumaJuan Felipe BautistaÎncă nu există evaluări
- Control y Regulación 1 III PeriodoDocument8 paginiControl y Regulación 1 III PeriodoCarlos Javier Almanza RodriguezÎncă nu există evaluări
- Influencia Del Medio Ambiente en Los OrganismoDocument7 paginiInfluencia Del Medio Ambiente en Los OrganismoPatrick HernandezÎncă nu există evaluări
- Neuroanatomia Del EstresDocument10 paginiNeuroanatomia Del EstresGianny PastorÎncă nu există evaluări
- Guia Didactica CastDocument18 paginiGuia Didactica CastJuan Pablo BlanchÎncă nu există evaluări
- Estres y Sistema InmuneDocument9 paginiEstres y Sistema InmuneJuan Fernando Matajudíos CarmonaÎncă nu există evaluări
- Psicobiología del estrés: homeostasis y respuesta al estrésDocument4 paginiPsicobiología del estrés: homeostasis y respuesta al estrésyessicaÎncă nu există evaluări
- EL ESTRÉS Y LA RELACIÓN CON PNEIDocument17 paginiEL ESTRÉS Y LA RELACIÓN CON PNEIpsicomerl2021Încă nu există evaluări
- Manual de Endocrino MitadDocument382 paginiManual de Endocrino MitadEvelyn ReinosoÎncă nu există evaluări
- Estres FawecDocument2 paginiEstres FawecHeidi Maggalí GarleppÎncă nu există evaluări
- Enfrenta TMB Cuestionamientos de Las Mente/cerebro. Frente A Este ProblemaDocument6 paginiEnfrenta TMB Cuestionamientos de Las Mente/cerebro. Frente A Este ProblemaSilvana CiccarelliÎncă nu există evaluări
- Trabajo Final BienestarDocument14 paginiTrabajo Final BienestarLucarno BantacoÎncă nu există evaluări
- Apego Cortisol y Estres ScieloDocument20 paginiApego Cortisol y Estres ScieloDaniela Alessandra DescalziÎncă nu există evaluări
- Problemática de Los Animales CallejerosDocument6 paginiProblemática de Los Animales CallejerosMaría de los ÁngelesÎncă nu există evaluări
- La Neuroquímica Del Estrés y El Papel de Los Péptidos Opioides PDFDocument9 paginiLa Neuroquímica Del Estrés y El Papel de Los Péptidos Opioides PDFRoller VizcarraÎncă nu există evaluări
- Principios Generales Sobre Estrés y Bienestar. Conductas EstereotipadasTEMA 19.-.Document28 paginiPrincipios Generales Sobre Estrés y Bienestar. Conductas EstereotipadasTEMA 19.-.Angélica Lozano CastroÎncă nu există evaluări
- Cronoterapia: principios y aplicacionesDocument6 paginiCronoterapia: principios y aplicacionesyinecologicÎncă nu există evaluări
- Ensayo de Bases Biologicas Del ComportamientoDocument6 paginiEnsayo de Bases Biologicas Del ComportamientoJose SilverioÎncă nu există evaluări
- Mecanismos epigenéticos de la depresiónDocument3 paginiMecanismos epigenéticos de la depresiónLluvia CrowleyÎncă nu există evaluări
- Alostatica 1Document75 paginiAlostatica 1Natalia SanchezÎncă nu există evaluări
- Psiconeuroinmunología: interacción mente-cuerpoDocument6 paginiPsiconeuroinmunología: interacción mente-cuerpoPaolaSerranoÎncă nu există evaluări
- Carga Alostatica 2Document89 paginiCarga Alostatica 2AMCArroyoÎncă nu există evaluări
- Ritmos BiológicosDocument4 paginiRitmos BiológicosMAGDALENA LOPEZÎncă nu există evaluări
- CronofarmacologiaDocument6 paginiCronofarmacologiajosuelugomendez97Încă nu există evaluări
- Respuesta Sistemática Ante El TraumaDocument5 paginiRespuesta Sistemática Ante El TraumaVanessa MendozaÎncă nu există evaluări
- Generalidades FisiopatologíaDocument71 paginiGeneralidades FisiopatologíaDulce patricia De la cruz gomezÎncă nu există evaluări
- Aportes y Futuro de La Psiconeuroinmunoendocrinología (Inmunoterapia) en La SaludDocument19 paginiAportes y Futuro de La Psiconeuroinmunoendocrinología (Inmunoterapia) en La SaludJair Monja Ramirez100% (1)
- El temperamento y su trama: Cómo los genes, la cultura, el tiempo y el azar inciden en nuestra personalidadDe la EverandEl temperamento y su trama: Cómo los genes, la cultura, el tiempo y el azar inciden en nuestra personalidadEvaluare: 5 din 5 stele5/5 (1)
- Convenio de Practicas 2015Document37 paginiConvenio de Practicas 2015Jean Carlos Motta OsmaÎncă nu există evaluări
- Convenio de Practicas 2015Document37 paginiConvenio de Practicas 2015Jean Carlos Motta OsmaÎncă nu există evaluări
- Proyecto ANH Temática 1Document1 paginăProyecto ANH Temática 1Jean Carlos Motta OsmaÎncă nu există evaluări
- Proyecto Mapa EcosistemasDocument1 paginăProyecto Mapa EcosistemasJean Carlos Motta OsmaÎncă nu există evaluări
- Spe 37856Document5 paginiSpe 37856Jean Carlos Motta OsmaÎncă nu există evaluări
- Fuentes Bibliograficas A ConsultarDocument1 paginăFuentes Bibliograficas A ConsultarJean Carlos Motta OsmaÎncă nu există evaluări
- Abanicos AluvialesDocument8 paginiAbanicos AluvialesJean Carlos Motta OsmaÎncă nu există evaluări
- Nimf 15Document13 paginiNimf 15milagros huayta matosÎncă nu există evaluări
- Asignacion #2 1-2021-2022Document3 paginiAsignacion #2 1-2021-2022Raimy Bisono GarciaÎncă nu există evaluări
- Frescura Del PescadoDocument21 paginiFrescura Del PescadodmontesdosÎncă nu există evaluări
- Sesión de Matemática 19 de OctubreDocument4 paginiSesión de Matemática 19 de OctubreMilagros CasasÎncă nu există evaluări
- Evaluacion Final - Escenario 8 - PRIMER BLOQUE-TEORICO-PRACTICO - VIRTUAL - SEMIÓTICA - (GRUPO B01)Document13 paginiEvaluacion Final - Escenario 8 - PRIMER BLOQUE-TEORICO-PRACTICO - VIRTUAL - SEMIÓTICA - (GRUPO B01)Nikolas RodriguezÎncă nu există evaluări
- Tarea1. .Document50 paginiTarea1. .Rene De La CruzÎncă nu există evaluări
- 1.sentencia de Casación 12482-2019-LIMA, Vinculada Con Las Multas Por Supuestas Omisiones de Pagos A Cuenta Del Impuesto A La RentaDocument11 pagini1.sentencia de Casación 12482-2019-LIMA, Vinculada Con Las Multas Por Supuestas Omisiones de Pagos A Cuenta Del Impuesto A La RentaShery Solis DurandÎncă nu există evaluări
- Prática 3 Tratamientos Térmicos y DurezaDocument13 paginiPrática 3 Tratamientos Térmicos y DurezaBryan Hernandez AlvaradoÎncă nu există evaluări
- Proyecto OficialDocument37 paginiProyecto OficialCristian CollaguazoÎncă nu există evaluări
- Boletin de La Papa, Vol 11, No. 12Document2 paginiBoletin de La Papa, Vol 11, No. 12Jorge Luis Alonso G.Încă nu există evaluări
- La Sistematica Base Del Conocimiento de La Biodiversidad 2007Document155 paginiLa Sistematica Base Del Conocimiento de La Biodiversidad 2007Cindy Castro GutierrezÎncă nu există evaluări
- Manual Del Usuario WixDocument6 paginiManual Del Usuario WixAndrés GarcíaÎncă nu există evaluări
- Practica No8 Reflejos en El Humano.-.Document14 paginiPractica No8 Reflejos en El Humano.-.Leion UchihaÎncă nu există evaluări
- Método completo de bateríaDocument10 paginiMétodo completo de bateríaMario Barrett100% (1)
- Manual de Organizacion CESPT PDFDocument213 paginiManual de Organizacion CESPT PDFJesus DomÎncă nu există evaluări
- InfografÍa Análisis de PuestoDocument2 paginiInfografÍa Análisis de PuestoYESICA MARISOL SANCHEZ SOSA100% (1)
- Publicidad EngañosaDocument2 paginiPublicidad EngañosaMilagro CepedaÎncă nu există evaluări
- Pseudomona AeruginosaDocument22 paginiPseudomona AeruginosaMiguel LCzÎncă nu există evaluări
- Pastores en crisis: 20 factores que dificultan el ministerioDocument6 paginiPastores en crisis: 20 factores que dificultan el ministerioGabriel GironÎncă nu există evaluări
- El rol del psicólogo: una ciencia para comprender y mejorar la conducta humanaDocument21 paginiEl rol del psicólogo: una ciencia para comprender y mejorar la conducta humanaYusmely Barcenas VillalbaÎncă nu există evaluări
- Startup e InnovacionDocument46 paginiStartup e InnovacionAdolfo GalindoÎncă nu există evaluări
- Lab 2Document17 paginiLab 2Patrick Hitedoshi Nakamura VillavicencioÎncă nu există evaluări
- Taller n.2 - El Dios Que AdoramosDocument4 paginiTaller n.2 - El Dios Que AdoramosOliviaÎncă nu există evaluări
- TAREA-2 Historia de La Ingeniería CivilDocument13 paginiTAREA-2 Historia de La Ingeniería CivilMICHELLE ISABEL TORRES LOPEZÎncă nu există evaluări
- Procesos de La Inspiración y EspiraciónDocument1 paginăProcesos de La Inspiración y Espiracióndepartamento GPS A&B nro 2Încă nu există evaluări
- Materia RedaccionDocument43 paginiMateria RedaccionJENIFFER MAGALY VARGAS SHIGUANGOÎncă nu există evaluări
- Tarea MvaDocument5 paginiTarea MvaAdamaris Molar VázquezÎncă nu există evaluări
- TIPOS DE ARGUMENTOS EjerciciosDocument1 paginăTIPOS DE ARGUMENTOS EjerciciosValeria salazar50% (2)
- La Gaceta - Agosto 2003 - Entre Letras y Voces Populares Hispanicas (Siglo XV Al XVII)Document32 paginiLa Gaceta - Agosto 2003 - Entre Letras y Voces Populares Hispanicas (Siglo XV Al XVII)Jeff HuckleberryÎncă nu există evaluări
- Enc 10 PARTES DE LA MISA IIDocument2 paginiEnc 10 PARTES DE LA MISA IIDelsy Marianela SOCUALAYA ACUÑAÎncă nu există evaluări