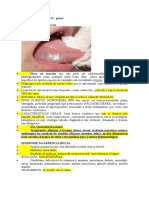
S-ar putea să vă placă și
- Abacate 2007Document8 paginiAbacate 2007Jennifer BeatrizÎncă nu există evaluări
- Biosintese Da Escopoletina e Escopolina em Mandioca Deterioração OkDocument43 paginiBiosintese Da Escopoletina e Escopolina em Mandioca Deterioração OkInaiara CasapulaÎncă nu există evaluări
- Bioquimica Aplicada Aula 3 Escurecimento EnzimaticoDocument26 paginiBioquimica Aplicada Aula 3 Escurecimento EnzimaticoLuiz PauloÎncă nu există evaluări
- Qualidade de Cultivares de Alface Produzidos em HidroponiaDocument5 paginiQualidade de Cultivares de Alface Produzidos em HidroponiaEstevan Butzke PyddÎncă nu există evaluări
- Processo 108 CaféDocument5 paginiProcesso 108 CaféPedro MarianoÎncă nu există evaluări
- Composição Química de Amêndoas Fermentadas de Cacau PDFDocument5 paginiComposição Química de Amêndoas Fermentadas de Cacau PDFLuiz CannalongaÎncă nu există evaluări
- BP 017Document22 paginiBP 017sorry welcomeÎncă nu există evaluări
- Slides Delfino PedroDocument25 paginiSlides Delfino Pedroamiel moraisÎncă nu există evaluări
- Extração, Purificaçao, Caract-LAP Gonçalo FigueiróDocument7 paginiExtração, Purificaçao, Caract-LAP Gonçalo FigueiróGonçalo AbrantesÎncă nu există evaluări
- 2017-MODIFICAÇÃO QUÍMICA DA PECTINA DO MELÃO CAIPIRA (Cucumis Melo VAR. ACIDULUS)Document7 pagini2017-MODIFICAÇÃO QUÍMICA DA PECTINA DO MELÃO CAIPIRA (Cucumis Melo VAR. ACIDULUS)Laboratorio y Control de Calidad Planta ProÎncă nu există evaluări
- Artigo - Produção de Enzimas - Cascas de Laranja e CocoDocument8 paginiArtigo - Produção de Enzimas - Cascas de Laranja e CocoAugusto SimasÎncă nu există evaluări
- Efeito Do Extrato Aquoso de Leucena Na Germinação E No Desenvolvimento Do Milho PDFDocument6 paginiEfeito Do Extrato Aquoso de Leucena Na Germinação E No Desenvolvimento Do Milho PDFMaria AndradeÎncă nu există evaluări
- Propolis Concentração PDFDocument7 paginiPropolis Concentração PDFSanta Bárbara do Leste Emater-MGÎncă nu există evaluări
- Qualidade de Grãos de Café Beneficiados emDocument7 paginiQualidade de Grãos de Café Beneficiados emJoão Carlos PinheiroÎncă nu există evaluări
- Estudo Da Reciclagem de Células Na Produção Biológica de EtanolDocument4 paginiEstudo Da Reciclagem de Células Na Produção Biológica de EtanolNandinha FloresÎncă nu există evaluări
- Biosintese Da Escopoletina e Escopolina em Mandioca Deterioração CertoDocument41 paginiBiosintese Da Escopoletina e Escopolina em Mandioca Deterioração CertoInaiara CasapulaÎncă nu există evaluări
- Composição e Benefícios Da Geoprópolis Da Abelha Melípona SubnitidaDocument24 paginiComposição e Benefícios Da Geoprópolis Da Abelha Melípona SubnitidaAmanda PeixotoÎncă nu există evaluări
- 1177 2674 1 PBDocument10 pagini1177 2674 1 PBJoseph PHÎncă nu există evaluări
- Constituintes Fenólicos e Atividade Antioxidante Da Geoprópolis de Duas Espécies de Abelhas Sem Ferrão AmazônicasDocument6 paginiConstituintes Fenólicos e Atividade Antioxidante Da Geoprópolis de Duas Espécies de Abelhas Sem Ferrão AmazônicasMZAÎncă nu există evaluări
- 223 869 1 PBDocument9 pagini223 869 1 PBRosiel Cabral PantojaÎncă nu există evaluări
- Dos Santos Et Al, 2014Document5 paginiDos Santos Et Al, 2014Vinicius MoreiraÎncă nu există evaluări
- Artigo Bea NovoDocument7 paginiArtigo Bea NovoDavi MagalhãesÎncă nu există evaluări
- PARTIÇÃO DE LIPASE DA AMÊNDOA DO PEQUI (Caryocar Brasiliense Camb.) EM SISTEMAS AQUOSOS BIFÁSICOS FORMADOS POR 2-PROPANOL + SULFATO DE AMÔNIO + ÁGUA.Document6 paginiPARTIÇÃO DE LIPASE DA AMÊNDOA DO PEQUI (Caryocar Brasiliense Camb.) EM SISTEMAS AQUOSOS BIFÁSICOS FORMADOS POR 2-PROPANOL + SULFATO DE AMÔNIO + ÁGUA.Mateus SantosÎncă nu există evaluări
- Relatório PráticaDocument12 paginiRelatório PráticabethfirmianoÎncă nu există evaluări
- Silagem de Milho e PupunhaDocument11 paginiSilagem de Milho e Pupunhaleandro mendesÎncă nu există evaluări
- 5677 Artigo 56419 1 10 20201119Document7 pagini5677 Artigo 56419 1 10 20201119giani.cavalcanteÎncă nu există evaluări
- Caracterização Da Polpa Farinácea e Semente Do JatobáDocument5 paginiCaracterização Da Polpa Farinácea e Semente Do JatobáLAURA FERREIRA DOS SANTOS RODRIGUESÎncă nu există evaluări
- Escurecimento Enzimático AlexDocument23 paginiEscurecimento Enzimático AlexdemidoverÎncă nu există evaluări
- UTILIZAÇÃO DE RESÍDUOS AGROINDUSTRIAIS PARA PRODUÇÃO DE AMILOGLUCOSIDASE POR Aspergillus AwamoriDocument10 paginiUTILIZAÇÃO DE RESÍDUOS AGROINDUSTRIAIS PARA PRODUÇÃO DE AMILOGLUCOSIDASE POR Aspergillus AwamoriLucas FigueiredoÎncă nu există evaluări
- Avaliação Da Qualidade de Polpa de Goiaba Paluma' Armazenada A - 20ºCDocument3 paginiAvaliação Da Qualidade de Polpa de Goiaba Paluma' Armazenada A - 20ºCCarlos Eduardo Robaina dos SantosÎncă nu există evaluări
- Arquivo 47960 1 10 20190923Document4 paginiArquivo 47960 1 10 20190923isaque clementeÎncă nu există evaluări
- Analise DPPHDocument8 paginiAnalise DPPHMárcia BayÎncă nu există evaluări
- Fermentado de UmbuDocument6 paginiFermentado de UmbuSolange FrazãoÎncă nu există evaluări
- Chá Verde Artigo CientificoDocument8 paginiChá Verde Artigo CientificoDiogoPC2015Încă nu există evaluări
- Capim ElefanteDocument6 paginiCapim ElefanteGiovana CazassaÎncă nu există evaluări
- Potencial Nutritivo de Polpas de MangostãoDocument9 paginiPotencial Nutritivo de Polpas de MangostãoClaraÎncă nu există evaluări
- Constituintes Voláteis de Própolis PiauienseDocument7 paginiConstituintes Voláteis de Própolis PiauienseMZAÎncă nu există evaluări
- Enzimas PectinolíticasDocument9 paginiEnzimas PectinolíticasDanielPiauÎncă nu există evaluări
- Preparation and Quality Jelly and Pineapple Compote "Pearl"Document5 paginiPreparation and Quality Jelly and Pineapple Compote "Pearl"PlayMusic BrÎncă nu există evaluări
- Artigo-Resultados Da Investigação-Tratamento de Agua Quiabo-114842-2-2-20220714Document14 paginiArtigo-Resultados Da Investigação-Tratamento de Agua Quiabo-114842-2-2-20220714Jose Pedro FranciscoÎncă nu există evaluări
- Cultivo de Melão Orgânico: Fosfatos Naturais Como Fontes Alternativas de FósforoDocument8 paginiCultivo de Melão Orgânico: Fosfatos Naturais Como Fontes Alternativas de FósforojandeilsonarrudaÎncă nu există evaluări
- Vinho - Figo Da Índia 4Document11 paginiVinho - Figo Da Índia 4AnaPaulaTormes0% (1)
- Alterações em AlimentosDocument13 paginiAlterações em AlimentosMariana RossoniÎncă nu există evaluări
- DownloadDocument6 paginiDownloadAvelino AugustoÎncă nu există evaluări
- Caracterização Físico-Química Da Polpa e Espuma de BacabaDocument5 paginiCaracterização Físico-Química Da Polpa e Espuma de BacabaValdenilson MoraisÎncă nu există evaluări
- 3207 - ESTUDO DA QUALIDADE DO CAFé SOMBREADO PRODUZIDO NO MACIçO DE BATURITé - CE - 3Document8 pagini3207 - ESTUDO DA QUALIDADE DO CAFé SOMBREADO PRODUZIDO NO MACIçO DE BATURITé - CE - 3Izaias AiresÎncă nu există evaluări
- RVq170420 A1Document8 paginiRVq170420 A1Heslen RodriguesÎncă nu există evaluări
- Artigo Cinetica de Degradacao de Geleiada de Morango Raiza PDFDocument6 paginiArtigo Cinetica de Degradacao de Geleiada de Morango Raiza PDFNatália CataniÎncă nu există evaluări
- Escurecimento EnzimáticoDocument18 paginiEscurecimento EnzimáticoKatia SouzaÎncă nu există evaluări
- Aula 5 - Alcaloides IsoquinolínicosDocument27 paginiAula 5 - Alcaloides IsoquinolínicosJoab Ana MenezesÎncă nu există evaluări
- Relatório Farmacognosia - PráticaDocument8 paginiRelatório Farmacognosia - PráticaDávila Kelly MaiaÎncă nu există evaluări
- Área: Engenharia de Alimentos: Caroline Marques, Angela R. de Almeida, Eliane D. Godoy DanesiDocument6 paginiÁrea: Engenharia de Alimentos: Caroline Marques, Angela R. de Almeida, Eliane D. Godoy DanesiClxssic XÎncă nu există evaluări
- Aplicacao Fruitone Frutos 1982Document3 paginiAplicacao Fruitone Frutos 1982Emanuel Maretto EffgenÎncă nu există evaluări
- FoulingDocument5 paginiFoulingLarissa Dos SantosÎncă nu există evaluări
- XilófagosDocument15 paginiXilófagoswelderÎncă nu există evaluări
- Pigmentação Do CamarãoDocument5 paginiPigmentação Do CamarãoCharlle LimaÎncă nu există evaluări
- Uso de Sistemas Aquosos Bifásicos Formados Por Peg + Fosfato de Sódio para Pré-Purificação de Proteases de Ora-Pro-Nóbis (Pereskia Aculeata Miller)Document6 paginiUso de Sistemas Aquosos Bifásicos Formados Por Peg + Fosfato de Sódio para Pré-Purificação de Proteases de Ora-Pro-Nóbis (Pereskia Aculeata Miller)Mateus SantosÎncă nu există evaluări
- Composição Químico-Bromatológica Da Casca de Café Tratada Com Amônia Anidra e Sulfeto de SódioDocument9 paginiComposição Químico-Bromatológica Da Casca de Café Tratada Com Amônia Anidra e Sulfeto de SódioHend GadowÎncă nu există evaluări
- Novas pesquisas em Ciências Biológicas e Biodiversidade: Volume 2De la EverandNovas pesquisas em Ciências Biológicas e Biodiversidade: Volume 2Încă nu există evaluări
- Ulva lactuca: Aspectos Químicos na Determinação da Atividade Antioxidante TotalDe la EverandUlva lactuca: Aspectos Químicos na Determinação da Atividade Antioxidante TotalÎncă nu există evaluări
- 09 Dentista - 20h - e - 40hDocument13 pagini09 Dentista - 20h - e - 40hRaíssa RibeiroÎncă nu există evaluări
- Centro Cultural Coreano - Livro - Portugues - 3Document152 paginiCentro Cultural Coreano - Livro - Portugues - 3Raíssa Ribeiro100% (2)
- Caso 1 Sangramento GengivalDocument15 paginiCaso 1 Sangramento GengivalRaíssa RibeiroÎncă nu există evaluări
- Bibliografia Recomendada OrtodontiaDocument1 paginăBibliografia Recomendada OrtodontiaRaíssa RibeiroÎncă nu există evaluări
- Grampos IacoDocument4 paginiGrampos IacoRaíssa RibeiroÎncă nu există evaluări
- Provão 2001 Gabarito - Prova OdontologiaDocument2 paginiProvão 2001 Gabarito - Prova OdontologiaRaíssa RibeiroÎncă nu există evaluări
- AmiloglicosilaseDocument7 paginiAmiloglicosilaseRaíssa RibeiroÎncă nu există evaluări
- APOSTILA EstomatoDocument28 paginiAPOSTILA EstomatoRaíssa RibeiroÎncă nu există evaluări
- Provão 1998 A 2003 Gabaritos Objetivos - Provas OdontologiaDocument6 paginiProvão 1998 A 2003 Gabaritos Objetivos - Provas OdontologiaRaíssa RibeiroÎncă nu există evaluări
- Provão 1999 Gabarito Subjetivo - Prova OdontologiaDocument4 paginiProvão 1999 Gabarito Subjetivo - Prova OdontologiaRaíssa RibeiroÎncă nu există evaluări
- Acidentes e ComplicaçõesDocument19 paginiAcidentes e ComplicaçõesRaíssa RibeiroÎncă nu există evaluări
- Hiperemia SeminarioDocument11 paginiHiperemia SeminarioRaíssa Ribeiro100% (1)
- Unidade III Disturbios CirculatoriosDocument100 paginiUnidade III Disturbios CirculatoriosRaíssa RibeiroÎncă nu există evaluări
- Editora BAGAI - Filosofia AfricanaDocument110 paginiEditora BAGAI - Filosofia Africanadomingos beula fortunato100% (1)
- Manual de Boas Práticas de FabricaçãoDocument8 paginiManual de Boas Práticas de FabricaçãoAlex AlmeidaÎncă nu există evaluări
- Henrique Murachco Gramatica Grega TeoriaDocument739 paginiHenrique Murachco Gramatica Grega Teorialatioricap0% (1)
- Tabela Cnu Pme Ma-1Document13 paginiTabela Cnu Pme Ma-1Sullivan SuadÎncă nu există evaluări
- TJ Ba Aula 1Document27 paginiTJ Ba Aula 1Lopes PublicidadeÎncă nu există evaluări
- Fichamento: MARTELOTTA. M, E. (Org.) - Manual de Linguística. São Paulo: Contexto, 2009, p.177 - 192Document2 paginiFichamento: MARTELOTTA. M, E. (Org.) - Manual de Linguística. São Paulo: Contexto, 2009, p.177 - 192Jason FelipeÎncă nu există evaluări
- As Microcervejarias No Brasil Atual - Eduardo MarcussoDocument171 paginiAs Microcervejarias No Brasil Atual - Eduardo MarcussoodairnettoÎncă nu există evaluări
- BriofitasDocument19 paginiBriofitasDalton Homisio100% (1)
- Avaliação 30 Dias NOVODocument4 paginiAvaliação 30 Dias NOVORodrigo GuterresÎncă nu există evaluări
- Dezembro LaranjaDocument7 paginiDezembro LaranjaLetíciaÎncă nu există evaluări
- Desenvolvimento de Um Dispositivo Detector de Vazamento de Gás Utilizando Arduino Como Interface de AutomaçãoDocument2 paginiDesenvolvimento de Um Dispositivo Detector de Vazamento de Gás Utilizando Arduino Como Interface de AutomaçãoLucas GodoiÎncă nu există evaluări
- MecFund Aula 08 - Oscilador Harmonico Amortecido PDFDocument57 paginiMecFund Aula 08 - Oscilador Harmonico Amortecido PDFLeonardoMaiaÎncă nu există evaluări
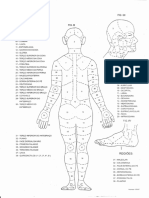
- Topografia Corpo HumanoDocument2 paginiTopografia Corpo Humanoestevaocanan100% (9)
- Lista de Exercícios (Força e Movimento I e II) - Física - Dinâmica A TermodinâmicaDocument6 paginiLista de Exercícios (Força e Movimento I e II) - Física - Dinâmica A TermodinâmicavagnerÎncă nu există evaluări
- Folheto Crisma 1Document2 paginiFolheto Crisma 1Murilo Vilas BoasÎncă nu există evaluări
- Manual Técnico Aparelho de Anestesia Takaoka Sat 500Document188 paginiManual Técnico Aparelho de Anestesia Takaoka Sat 500CENTRO OESTE100% (1)
- Transmissor Com Ba1404 e 2sc2458Document8 paginiTransmissor Com Ba1404 e 2sc2458carrlosalbertodossantossilvaÎncă nu există evaluări
- Iracema - Adaptação Teatral (José de Alencar)Document5 paginiIracema - Adaptação Teatral (José de Alencar)Zeca RolandÎncă nu există evaluări
- GREGOLIN, Remontemos de Foucault A Spinoza-PecheuxDocument11 paginiGREGOLIN, Remontemos de Foucault A Spinoza-Pecheuxraquel_rybandtÎncă nu există evaluări
- EDITAL PRODIC-UNEAL No 030-2020 - TURMA 6 - 2021Document21 paginiEDITAL PRODIC-UNEAL No 030-2020 - TURMA 6 - 2021jorgeÎncă nu există evaluări
- Arquitectura de Computadores PDFDocument99 paginiArquitectura de Computadores PDFpggsÎncă nu există evaluări
- 73 78Document6 pagini73 78Thiago AguiarÎncă nu există evaluări
- A Psicoterapia Breve - Abordagem TCCDocument19 paginiA Psicoterapia Breve - Abordagem TCCAdriana AraldiÎncă nu există evaluări
- Nery & Beck: Encordoando Histórias Do TênisDocument80 paginiNery & Beck: Encordoando Histórias Do TênissilvioÎncă nu există evaluări
- Historia e Geografia de GoiasDocument103 paginiHistoria e Geografia de GoiasMichel Merelles100% (2)
- 3-RenascimentoCultural e CientificoDocument21 pagini3-RenascimentoCultural e CientificoPaulo AndradeÎncă nu există evaluări
- Referencial de Competências-Chave NS STCDocument21 paginiReferencial de Competências-Chave NS STCnuno.jose.duarte100% (1)
- Questoes Revisao Histologia BasicaDocument20 paginiQuestoes Revisao Histologia BasicaAlessandra IngridÎncă nu există evaluări
- Vida Saudável PDFDocument4 paginiVida Saudável PDFKAUÃ GOMES DIASÎncă nu există evaluări
- Asma e Rinite Ocupacionais 2010Document6 paginiAsma e Rinite Ocupacionais 2010Li LiuÎncă nu există evaluări