S-ar putea să vă placă și
- Ecología Microbiana y Métodos de EstudioDocument6 paginiEcología Microbiana y Métodos de EstudioVALERIA LATORRE SUCNIERÎncă nu există evaluări
- Proyecto Investigativo - Ecosistema.Document7 paginiProyecto Investigativo - Ecosistema.jessica loorÎncă nu există evaluări
- Informe Final .. Ecosistema TerresteDocument7 paginiInforme Final .. Ecosistema Terrestejessica loorÎncă nu există evaluări
- PW Eco Microbiana 1 TEODocument13 paginiPW Eco Microbiana 1 TEOWILFREDO CHAVEZ HUAMANÎncă nu există evaluări
- Alvarez 2005 - La Descomposicion de Materia Organica en HumedalesDocument13 paginiAlvarez 2005 - La Descomposicion de Materia Organica en HumedalesScientist1718Încă nu există evaluări
- Monografia EcosistemasDocument17 paginiMonografia EcosistemasCassiopea Laura Ponce100% (1)
- Examen BiotecnologíaDocument3 paginiExamen BiotecnologíaJose Medina FelixÎncă nu există evaluări
- Investigación AmbientalDocument14 paginiInvestigación Ambientaljulio pineda urbinaÎncă nu există evaluări
- Biotecnologia 2Document72 paginiBiotecnologia 2alxcuchiaÎncă nu există evaluări
- Arbol Filogenetico de La VidaDocument11 paginiArbol Filogenetico de La VidaKevin MolinaÎncă nu există evaluări
- Informe Sobre Ecologia MicrobianaDocument15 paginiInforme Sobre Ecologia MicrobianaLuis SanchezÎncă nu există evaluări
- Ecologia MicrobianaDocument3 paginiEcologia MicrobianaYexon Mondragon SalvadorÎncă nu există evaluări
- 12 Asociaciones MicrobianasDocument16 pagini12 Asociaciones MicrobianasJoseph Crow SalazarÎncă nu există evaluări
- Ciclos BiogeoquímicosDocument52 paginiCiclos BiogeoquímicosJean Piero Campoverde CunyaÎncă nu există evaluări
- BiomarcadoresDocument11 paginiBiomarcadoresJhonatan RS0% (1)
- Una Microscópica Mirada A Los Suelos de Los Ecosistemas TropicalesDocument13 paginiUna Microscópica Mirada A Los Suelos de Los Ecosistemas TropicalesCarmen Beatriz DesantiagoÎncă nu există evaluări
- Ecologia MicrobianaDocument5 paginiEcologia MicrobianaYaxi FariasÎncă nu există evaluări
- Materiales T.P N º 1 AmbienteDocument4 paginiMateriales T.P N º 1 AmbienteBeatriz SanchezÎncă nu există evaluări
- Ensayo EsenarionaturalDocument7 paginiEnsayo EsenarionaturalCelso VazquezÎncă nu există evaluări
- Garcia y Bello 2004Document2 paginiGarcia y Bello 2004omarÎncă nu există evaluări
- Rol Ecologico HongosDocument25 paginiRol Ecologico HongosErick TsujiÎncă nu există evaluări
- Ecologia Microbiana DARWIN CHACHADocument20 paginiEcologia Microbiana DARWIN CHACHAJahirÎncă nu există evaluări
- Clase 1. Microbiología Ambiental 2019Document23 paginiClase 1. Microbiología Ambiental 2019MARIA INTRIAGO100% (1)
- EcologíaDocument6 paginiEcologíaKaren SemecoÎncă nu există evaluări
- Texto Nro 2 MicrobiologíaDocument2 paginiTexto Nro 2 Microbiologíaricardematias555Încă nu există evaluări
- ANÁLISIS DEL TEMA Video 2 Microbiologia AmbientalDocument3 paginiANÁLISIS DEL TEMA Video 2 Microbiologia AmbientalVanessa LópezÎncă nu există evaluări
- BIOÉTICA Y EcologiaDocument10 paginiBIOÉTICA Y EcologiaGeraldine Guerrero100% (1)
- Ensayo - Ecologia y Medio Ambiente (Monica Vela)Document7 paginiEnsayo - Ecologia y Medio Ambiente (Monica Vela)Mónica VelaÎncă nu există evaluări
- Relacion de La Bioquimica Con Ingenieria AmbientalDocument8 paginiRelacion de La Bioquimica Con Ingenieria AmbientalRosario Alonzo Alva0% (3)
- COMTAMINACIONAMBIENTAL MICROORGANISMOS Proyecto de KarinaDocument7 paginiCOMTAMINACIONAMBIENTAL MICROORGANISMOS Proyecto de KarinaJakelin G. Espinoza SandovalÎncă nu există evaluări
- Ecologia Microbiana IntroDocument19 paginiEcologia Microbiana Introisauraampuero9481100% (1)
- Reconocimiento Microbiologia de SueloDocument13 paginiReconocimiento Microbiologia de SueloAnaNellyMosqueraÎncă nu există evaluări
- TP 11 - 12 (2020)Document18 paginiTP 11 - 12 (2020)marcelinÎncă nu există evaluări
- Algo Muy Complejo Sobre Los Grupos FuncionalesDocument270 paginiAlgo Muy Complejo Sobre Los Grupos FuncionalesAmado MartinezÎncă nu există evaluări
- Taller II de Ecología BásicaDocument39 paginiTaller II de Ecología BásicadejomiÎncă nu există evaluări
- MICROORGANISMS AND CC JUN2019 CAVICCHIOLI - En.es-1-14Document14 paginiMICROORGANISMS AND CC JUN2019 CAVICCHIOLI - En.es-1-14Teresa MuchaÎncă nu există evaluări
- Ensayo2U6 Equipo6Document10 paginiEnsayo2U6 Equipo6Angie AcostaÎncă nu există evaluări
- 1.4 Aplicaciones de Microorganismos BioremediacionDocument8 pagini1.4 Aplicaciones de Microorganismos BioremediacionALeDuckBoanergesÎncă nu există evaluări
- The Emergence of Regularity and Variability in MarDocument13 paginiThe Emergence of Regularity and Variability in Marbiany paola celis caicedoÎncă nu există evaluări
- Práctica N°9 - Propiedades Biológicas Del SueloDocument21 paginiPráctica N°9 - Propiedades Biológicas Del SueloDaniela MonsalveÎncă nu există evaluări
- S1.-Ecologìa Microbiana y Microbiologìa AmbientalDocument20 paginiS1.-Ecologìa Microbiana y Microbiologìa AmbientalLeinZLT0% (2)
- Ecología Atómica - Los Ecosistemas Explicados A Partir de Los Átomos Que Los Forman - UABDivulga Barcelona Investigación e InnovaciónDocument3 paginiEcología Atómica - Los Ecosistemas Explicados A Partir de Los Átomos Que Los Forman - UABDivulga Barcelona Investigación e Innovaciónruthmaricieloquispeventura372Încă nu există evaluări
- Instructivo de Practica Semana 11Document2 paginiInstructivo de Practica Semana 11luis carlos marreros abantoÎncă nu există evaluări
- Microbiología y Medio AmbienteDocument6 paginiMicrobiología y Medio AmbienteAlexÎncă nu există evaluări
- Actividad 1. ECOLOGÍADocument11 paginiActividad 1. ECOLOGÍAWilson José Manchego PerdomoÎncă nu există evaluări
- Ecología y Ciencias AfinesDocument6 paginiEcología y Ciencias AfinesSalvbert GonzalezÎncă nu există evaluări
- Barreros Santiago-Resumen Biotecnología AgrícolaDocument34 paginiBarreros Santiago-Resumen Biotecnología AgrícolaSantiStalinÎncă nu există evaluări
- IA G1 B20 Torres Ron Perla CarolinaDocument18 paginiIA G1 B20 Torres Ron Perla CarolinaPerla RonÎncă nu există evaluări
- PA2 Hugo OrdoñezDocument3 paginiPA2 Hugo OrdoñezHugo Alberto Ordoñez ValdezÎncă nu există evaluări
- Microbiología AmbientalDocument8 paginiMicrobiología AmbientalSayan Garcia RuizÎncă nu există evaluări
- Trabajo Sobre La EcologiaDocument3 paginiTrabajo Sobre La EcologiaAnyalery Torres ToroÎncă nu există evaluări
- Conservación AmbientalDocument8 paginiConservación AmbientalJUAN LUIS RUIZÎncă nu există evaluări
- Ecología y MicrobiologíaDocument21 paginiEcología y MicrobiologíaFlor Yépez JimenezÎncă nu există evaluări
- SESION 5.pptx CICLOS BIOGEOQUIMICOSDocument87 paginiSESION 5.pptx CICLOS BIOGEOQUIMICOSXimenaÎncă nu există evaluări
- Aportes de la biología del suelo a la agroecologíaDe la EverandAportes de la biología del suelo a la agroecologíaEvaluare: 5 din 5 stele5/5 (1)
- Pgma U2 Ea HNVCDocument10 paginiPgma U2 Ea HNVCHugo VeraÎncă nu există evaluări
- Disciplinas Que Se Relacionan Con La EcologíaDocument6 paginiDisciplinas Que Se Relacionan Con La EcologíaGonzaloRamírezÎncă nu există evaluări
- 34 Ecolmicrob Actsem 34Document4 pagini34 Ecolmicrob Actsem 34Hana BaltazarÎncă nu există evaluări
- Ecología y Medio AmbienteDocument6 paginiEcología y Medio AmbienteMononokeHimeÎncă nu există evaluări
- Fragilidad Expuesta. Organismos Vulnerables al Cambio Climático.De la EverandFragilidad Expuesta. Organismos Vulnerables al Cambio Climático.Încă nu există evaluări
- Gilberto YanaricoDocument12 paginiGilberto YanaricoJulio CamelÎncă nu există evaluări
- Maestria en Ecologia AplicadaDocument1 paginăMaestria en Ecologia AplicadaJulio CamelÎncă nu există evaluări
- Asimilación de Cianuro Por BacteriasDocument6 paginiAsimilación de Cianuro Por BacteriasJulio CamelÎncă nu există evaluări
- Diego Guerrero - Resumen de El CapitalDocument81 paginiDiego Guerrero - Resumen de El CapitalBryan Alexander Valladares PeñaililloÎncă nu există evaluări
- Tomas CayoDocument11 paginiTomas CayoJulio CamelÎncă nu există evaluări
- Balance de Agua ActualizadoDocument2 paginiBalance de Agua ActualizadoJulio CamelÎncă nu există evaluări
- Plantilla Nombramiento JuridicaDocument1 paginăPlantilla Nombramiento JuridicaJulio CamelÎncă nu există evaluări
- Oxidación Microbiana de Minerales de Oro y Biolixiviación de OroDocument7 paginiOxidación Microbiana de Minerales de Oro y Biolixiviación de OroJulio CamelÎncă nu există evaluări
- Teorias de LiderazgoDocument24 paginiTeorias de LiderazgoDani Elías VásquezÎncă nu există evaluări
- Informe Bienal Sobre Realidad UniversitariaDocument134 paginiInforme Bienal Sobre Realidad UniversitariaabogadolaraÎncă nu există evaluări
- Liderazgo COMUNICACIONDocument8 paginiLiderazgo COMUNICACIONJulio CamelÎncă nu există evaluări
- Clase Uni 2017Document40 paginiClase Uni 2017victorÎncă nu există evaluări
- Alta Dirección LiderazgoDocument5 paginiAlta Dirección LiderazgoJulio CamelÎncă nu există evaluări
- Luis LoaizaDocument27 paginiLuis LoaizaJulio CamelÎncă nu există evaluări
- Joseph ChancasanampaDocument30 paginiJoseph ChancasanampaJulio Camel100% (1)
- Jorge Soto YenDocument23 paginiJorge Soto YenJulio CamelÎncă nu există evaluări
- Tomas CayoDocument11 paginiTomas CayoJulio CamelÎncă nu există evaluări
- Erika Calmell Del Solar PDFDocument20 paginiErika Calmell Del Solar PDFJulio CamelÎncă nu există evaluări
- Leoncio SevillaDocument17 paginiLeoncio SevillaJulio CamelÎncă nu există evaluări
- Marco Antonio FloresDocument28 paginiMarco Antonio FloresJulio CamelÎncă nu există evaluări
- Gino Bianchi MosqueraDocument22 paginiGino Bianchi MosqueraJulio CamelÎncă nu există evaluări
- Deicy SanchezDocument25 paginiDeicy SanchezJulio CamelÎncă nu există evaluări
- Modelacion Geologica Del Deposito - II ADocument165 paginiModelacion Geologica Del Deposito - II AJulio CamelÎncă nu există evaluări
- Elizabeth MezaDocument17 paginiElizabeth MezaJulio CamelÎncă nu există evaluări
- Joseph ChancasanampaDocument30 paginiJoseph ChancasanampaJulio Camel100% (1)
- Jose ConchaDocument19 paginiJose ConchaJulio CamelÎncă nu există evaluări
- Tomas CayoDocument11 paginiTomas CayoJulio CamelÎncă nu există evaluări
- Gustavo de La TorreDocument26 paginiGustavo de La TorreJulio CamelÎncă nu există evaluări
- Daniel García RamosDocument53 paginiDaniel García RamosJulio CamelÎncă nu există evaluări
- Trabajo Modulo I - InflaciónDocument16 paginiTrabajo Modulo I - InflaciónCarlos PeÎncă nu există evaluări
- SEMANA 14 Completamiento de TextosDocument8 paginiSEMANA 14 Completamiento de TextosEmerson Junior Ramos CotrinaÎncă nu există evaluări
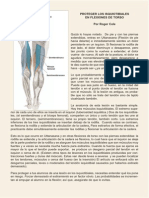
- Proteger Los IsquiotibialesDocument4 paginiProteger Los IsquiotibialesYogayeducacionÎncă nu există evaluări
- Sanidad Interior-Tesis FinalDocument54 paginiSanidad Interior-Tesis FinalIsrael Maravilla Arevalo100% (7)
- Guía de Aprendizaje #2 - 41311617Document9 paginiGuía de Aprendizaje #2 - 41311617rosanaÎncă nu există evaluări
- RESUMEN CUARTA Y QUINTA SEMANA (Imbestigacion Criminal) .Document3 paginiRESUMEN CUARTA Y QUINTA SEMANA (Imbestigacion Criminal) .Nilson Torres DiazÎncă nu există evaluări
- Planeacion Agregada Produccion IIDocument17 paginiPlaneacion Agregada Produccion IIGarcia VladimirÎncă nu există evaluări
- PRONABEC - ENP - PRONABEC4 - Simulacro PDFDocument17 paginiPRONABEC - ENP - PRONABEC4 - Simulacro PDFYeimis Francisco SaavedraÎncă nu există evaluări
- Planeación - TerpelDocument2 paginiPlaneación - Terpelcarolinaariza1089Încă nu există evaluări
- Planilla de Derecho A Matrícula OCRE-ULADocument1 paginăPlanilla de Derecho A Matrícula OCRE-ULAAlen UzcáteguiÎncă nu există evaluări
- Prisión VerdeDocument4 paginiPrisión VerdeMariela OsortoÎncă nu există evaluări
- Publicación - Anuncio - Bases Generales Selección Personal Funcionario y Laboral Interino Ayuntamiento UbriqueDocument6 paginiPublicación - Anuncio - Bases Generales Selección Personal Funcionario y Laboral Interino Ayuntamiento UbriqueJose Manuel Gómez LópezÎncă nu există evaluări
- Clase 3 Transporte de Contenedores A GranelDocument23 paginiClase 3 Transporte de Contenedores A GranelRonald CortezÎncă nu există evaluări
- Kínder Música Clase 6 10-05-2021Document5 paginiKínder Música Clase 6 10-05-2021Claudia HernándezÎncă nu există evaluări
- ViguetasDocument8 paginiViguetasBrunitoValeraÎncă nu există evaluări
- Ecoturismo en AmazoniaDocument18 paginiEcoturismo en AmazoniaCarolina SánchezÎncă nu există evaluări
- Planificaciones Dua LenguajeDocument6 paginiPlanificaciones Dua LenguajeClaudia Martinez100% (1)
- Matriz Productiva.Document14 paginiMatriz Productiva.Dennis GuamanÎncă nu există evaluări
- Actividades Primarias Del HombreDocument2 paginiActividades Primarias Del HombreFernando Suge Knight AmaruÎncă nu există evaluări
- ContratoDocument3 paginiContratoceleste tejera mezaÎncă nu există evaluări
- Ta-1 - Critica 1 Trabajo de Investigacion TechoDocument1 paginăTa-1 - Critica 1 Trabajo de Investigacion TechoGianella LunaÎncă nu există evaluări
- Revista de ArteDocument8 paginiRevista de ArteGustavo RadiceÎncă nu există evaluări
- Tesis Residuos SolidosDocument107 paginiTesis Residuos SolidosAlexander Calderon AguilarÎncă nu există evaluări
- Renault Nissan PDFDocument23 paginiRenault Nissan PDFtyronepÎncă nu există evaluări
- Sistema Constructivo DrywallDocument19 paginiSistema Constructivo DrywallBryan TlvÎncă nu există evaluări
- NombramientoDocument1 paginăNombramientoAndres PeñalozaÎncă nu există evaluări
- La Ortografia Grupo Mancha NegraDocument6 paginiLa Ortografia Grupo Mancha NegraBilbo BolsónÎncă nu există evaluări
- Paleogeografía de La Era TerciariaDocument12 paginiPaleogeografía de La Era TerciariaNataly SerranoÎncă nu există evaluări
- Trabajo Juicio Por JuradosDocument26 paginiTrabajo Juicio Por JuradosGina Camila MoroniÎncă nu există evaluări
- La Salvación Selectiva o PredestinaciónDocument5 paginiLa Salvación Selectiva o PredestinacióndistritosamariaÎncă nu există evaluări