S-ar putea să vă placă și
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Contractia MuscularaDocument26 paginiContractia Muscularadidu91100% (1)
- BFZ Contractia Musculara 2018Document61 paginiBFZ Contractia Musculara 2018Ashraf AlkalbaniÎncă nu există evaluări
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- Miologie GeneralaDocument4 paginiMiologie GeneralaIoana Ruxandra AnghelÎncă nu există evaluări
- Caracteristici Morfofunctionale Ale Aparatului LocomotorDocument4 paginiCaracteristici Morfofunctionale Ale Aparatului LocomotorSilviu LipanÎncă nu există evaluări
- Leziunile de Nervi Periferici Ghid de Diagnostic Si TratamentDocument85 paginiLeziunile de Nervi Periferici Ghid de Diagnostic Si Tratamentizo_katenaÎncă nu există evaluări
- Reflex eDocument10 paginiReflex eMadalina MarinÎncă nu există evaluări
- Suport de Curs Anatomie BuldușDocument65 paginiSuport de Curs Anatomie BuldușToma Bogdan-Laurentiu100% (2)
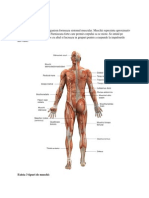
- Sistemul MuscularDocument17 paginiSistemul Muscularpoetu0Încă nu există evaluări
- 13.3. TermoterapiaDocument31 pagini13.3. TermoterapiaSorin LuchianÎncă nu există evaluări
- Articulatiile Coloanei Vertebrale - ANATOMIE - OdpDocument17 paginiArticulatiile Coloanei Vertebrale - ANATOMIE - OdpLazar Mihail100% (2)
- Coloana Lombara Curs 8Document6 paginiColoana Lombara Curs 8Gligan Alin-CristianÎncă nu există evaluări
- Principiile Morale Ale KinetoterapeutuluiDocument16 paginiPrincipiile Morale Ale KinetoterapeutuluiDaniela Paduraru100% (1)
- Nervul FemuralDocument1 paginăNervul FemuralstancuelenaÎncă nu există evaluări
- Curs Neuro 2 2010Document40 paginiCurs Neuro 2 2010Ciprian Cozmuleasa100% (3)
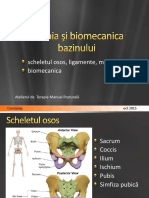
- Anatomia Si Biomecanica BazinuluiDocument21 paginiAnatomia Si Biomecanica BazinuluiAlexandru NechiforÎncă nu există evaluări
- Planse Articulatii+MembreDocument19 paginiPlanse Articulatii+MembreMarian DraganÎncă nu există evaluări
- 13.neuron Motor PerifericDocument16 pagini13.neuron Motor PerifericCristina AlexaÎncă nu există evaluări
- Tabele EvaluareDocument12 paginiTabele EvaluareDuta DanielaÎncă nu există evaluări
- NeurologieDocument48 paginiNeurologieMelisaÎncă nu există evaluări
- Articulatii - LPDocument7 paginiArticulatii - LPclaudia_gheneaÎncă nu există evaluări
- Seminar 2 Tehnici de Manevrare A BolnavuluiDocument2 paginiSeminar 2 Tehnici de Manevrare A BolnavuluiJst DanezuÎncă nu există evaluări
- Biomecanica MuscularaDocument30 paginiBiomecanica MuscularaBaciu Diana100% (2)
- Indicatii Si Contraindicatii Ale MasajuluiDocument2 paginiIndicatii Si Contraindicatii Ale MasajuluiDraghici Valentin100% (1)
- Elemente de FizioterapieDocument13 paginiElemente de FizioterapieCris TinaÎncă nu există evaluări
- Maduva SpinariiDocument12 paginiMaduva Spinariieniko1990100% (2)
- Curs - Evaluarea Pacientului Reumatic - 2020Document27 paginiCurs - Evaluarea Pacientului Reumatic - 2020Stefan AlexandruÎncă nu există evaluări
- BAZEDocument5 paginiBAZEPasvantu DanielÎncă nu există evaluări
- Motilitate ReflexeDocument9 paginiMotilitate ReflexeNorbert StenczelÎncă nu există evaluări
- Osteologie Si ArtrologieDocument51 paginiOsteologie Si ArtrologieCristina IoanaÎncă nu există evaluări
- Str. NeuronuluiDocument7 paginiStr. NeuronuluiLehaci Adelina100% (1)
- Biochima MuschilorDocument7 paginiBiochima MuschilorstikssÎncă nu există evaluări
- Anatomia Piciorului c1Document21 paginiAnatomia Piciorului c1CiureaDanaÎncă nu există evaluări
- Avc KinetoDocument80 paginiAvc KinetoRashid MihaiÎncă nu există evaluări
- Afectiuni Musculare - Semne Si Simptome - Curs 1Document44 paginiAfectiuni Musculare - Semne Si Simptome - Curs 1paulpop100% (2)
- Artera Femurala Plexul Lombar Plexul SacratDocument17 paginiArtera Femurala Plexul Lombar Plexul Sacratdanutsa89Încă nu există evaluări
- Curentii de Inalta FrecventaDocument26 paginiCurentii de Inalta FrecventaroxanabuciuÎncă nu există evaluări
- Kinesiologie 02Document8 paginiKinesiologie 02Contratov Lucian AndreiÎncă nu există evaluări
- Sistemul LimfaticDocument54 paginiSistemul LimfaticIonela ZubcoÎncă nu există evaluări
- Tesut NervosDocument6 paginiTesut NervosAnonymous pPacR7ydBÎncă nu există evaluări
- Muschi IDocument5 paginiMuschi IAlex CosminÎncă nu există evaluări
- Principiile Medicinei Traditionale Chineze Aplicate in Educatie Fizica Si KinetoterapieDocument1 paginăPrincipiile Medicinei Traditionale Chineze Aplicate in Educatie Fizica Si Kinetoterapiedoina13Încă nu există evaluări
- Unit 1GENERALITĂȚI PRIVIND ECHIPAMENTE, INSTALAȚII ȘI APARATE UTILIZATE ÎN KINETOTERAPIEDocument17 paginiUnit 1GENERALITĂȚI PRIVIND ECHIPAMENTE, INSTALAȚII ȘI APARATE UTILIZATE ÎN KINETOTERAPIEAndrei Crs100% (1)
- Fiziopatologia MuschiuluiDocument11 paginiFiziopatologia MuschiuluiSimona Boaru50% (2)
- Curs Biomecanica PDFDocument24 paginiCurs Biomecanica PDFFotea GeorgianaÎncă nu există evaluări
- 2 Kinetoterapie MARCUDocument51 pagini2 Kinetoterapie MARCUCosmin CezarÎncă nu există evaluări
- InimaDocument5 paginiInimaPai TavyÎncă nu există evaluări
- Plexurile NervoaseDocument4 paginiPlexurile NervoaseCristian GrecuÎncă nu există evaluări
- Gabat 1Document165 paginiGabat 1andrei_costachiÎncă nu există evaluări
- Fiziologia MuschilorDocument15 paginiFiziologia Muschilorpstefan18100% (1)
- Anatomia Maduvei SpinariiDocument8 paginiAnatomia Maduvei Spinariilala_girlÎncă nu există evaluări
- 9.sindromul MiofascialDocument5 pagini9.sindromul Miofascialdanyela_xp3071Încă nu există evaluări
- Biofizica Contractiei MusculareDocument6 paginiBiofizica Contractiei MusculareAndrone CameliaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- Biofizica Contractiei Musculare Sistemul Muscular PDFDocument1 paginăBiofizica Contractiei Musculare Sistemul Muscular PDFandamuresan3Încă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Structura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDocument2 paginiStructura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDaniela IstratiÎncă nu există evaluări
- Curs 5 BiofizicaDocument9 paginiCurs 5 BiofizicacurteanancaÎncă nu există evaluări
- Oscar Si Tanti RozDocument74 paginiOscar Si Tanti Rozakexandra83% (6)
- Sfaturi in Anul 2 2018-2019Document8 paginiSfaturi in Anul 2 2018-2019costica16100% (1)
- 05 Bioelectricitate 2017-12-08Document15 pagini05 Bioelectricitate 2017-12-08costica16Încă nu există evaluări
- AcidobazicDocument7 paginiAcidobazicAndrei MaricaÎncă nu există evaluări
- PoliticaDocument2 paginiPoliticacostica16Încă nu există evaluări
- 04 Membrane - 2017 12 08Document16 pagini04 Membrane - 2017 12 08costica16Încă nu există evaluări
- 03 - Sisteme - Disperse 2017 12 08Document13 pagini03 - Sisteme - Disperse 2017 12 08costica16Încă nu există evaluări
- Proteoglicani: Proteina Glicozaminoglican Trizaharid de LegãturãDocument7 paginiProteoglicani: Proteina Glicozaminoglican Trizaharid de Legãturãcostica16Încă nu există evaluări
- 02 - Apa 2017 12 04Document9 pagini02 - Apa 2017 12 04costica16Încă nu există evaluări
- 01 - Termodinamica 2017 12 04Document9 pagini01 - Termodinamica 2017 12 04costica16Încă nu există evaluări
- Pasta de Ardei Copti Cu Usturoi La Borcan Reteta de LutenitaDocument10 paginiPasta de Ardei Copti Cu Usturoi La Borcan Reteta de Lutenitacostica16Încă nu există evaluări
- Efectul PlaceboDocument3 paginiEfectul Placebocostica16Încă nu există evaluări
- Siruri - Calculul Limitelor Sirurilor Date Prin Termenul GeneralDocument15 paginiSiruri - Calculul Limitelor Sirurilor Date Prin Termenul GeneralBogdan Pisai75% (16)
- Compusi CarboniliciDocument47 paginiCompusi Carbonilicicostica16Încă nu există evaluări
- Simulare MG 2014 PDFDocument7 paginiSimulare MG 2014 PDFSebiTenorulÎncă nu există evaluări
- Ludwig Van BeethovenDocument4 paginiLudwig Van Beethovencostica16100% (1)
- Chimie OrganicaDocument10 paginiChimie OrganicaPavelMihaelaÎncă nu există evaluări
- Presstern Memorator Chimie 2 OrganicaDocument22 paginiPresstern Memorator Chimie 2 OrganicapylotÎncă nu există evaluări
- Petrol UlDocument3 paginiPetrol Ulcostica16Încă nu există evaluări
- Sinteza - Sistemul MuscularDocument11 paginiSinteza - Sistemul Muscularlesnoland100% (1)
- Proiect PRAGA Geogra Cls 10Document3 paginiProiect PRAGA Geogra Cls 10costica16Încă nu există evaluări
- Sinteza-Sistemul Osos Si ArticulatiileDocument10 paginiSinteza-Sistemul Osos Si ArticulatiileIon CovaliÎncă nu există evaluări
- Probleme Reprrezzolvate Clasa A IxDocument26 paginiProbleme Reprrezzolvate Clasa A IxSalcianu IonutÎncă nu există evaluări
- Panouri RadianteDocument7 paginiPanouri Radiantecostica16Încă nu există evaluări
- Probleme 10 E MatriciDocument4 paginiProbleme 10 E MatricishownsharkÎncă nu există evaluări
- Johannes BrahmsDocument9 paginiJohannes Brahmscostica16Încă nu există evaluări
- Plan de AfaceriDocument3 paginiPlan de Afacericostica16Încă nu există evaluări
- Johannes BrahmsDocument9 paginiJohannes Brahmscostica16Încă nu există evaluări
- I Razboi MondialDocument5 paginiI Razboi Mondialcostica16Încă nu există evaluări