S-ar putea să vă placă și
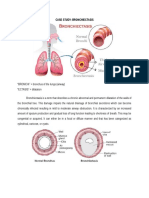
- BronchiectasisDocument21 paginiBronchiectasisMuhammad Iyhan NuriansyahÎncă nu există evaluări
- Medical Progress: Review ArticleDocument11 paginiMedical Progress: Review ArticlemayaÎncă nu există evaluări
- Bronchiectasis: DR Budi Enoch SPPDDocument21 paginiBronchiectasis: DR Budi Enoch SPPDVito CuyÎncă nu există evaluări
- Main Image ©muratseyit/istockphotoDocument10 paginiMain Image ©muratseyit/istockphotoade_liaÎncă nu există evaluări
- Bronquiolite DiagnosticoDocument14 paginiBronquiolite DiagnosticoFrederico PóvoaÎncă nu există evaluări
- Bronchiectasis OkDocument60 paginiBronchiectasis OkImmanuelÎncă nu există evaluări
- Bronchiectasis: Yuping GuoDocument75 paginiBronchiectasis: Yuping GuoAmeliaÎncă nu există evaluări
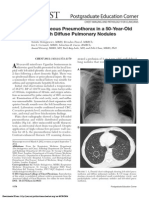
- Chest X-Ray Manifestations of PneumoniaDocument3 paginiChest X-Ray Manifestations of PneumoniaVishesh SoodÎncă nu există evaluări
- 3 BronchiectasisDocument29 pagini3 BronchiectasisЕвгений ХанькоÎncă nu există evaluări
- Diagnostic Evaluation of BronchiectasisDocument16 paginiDiagnostic Evaluation of BronchiectasisAinun Jariah MuliadiÎncă nu există evaluări
- Grenier 2009Document18 paginiGrenier 2009PabloÎncă nu există evaluări
- Challenges in Pulmonary Fibrosis ? 3: Cystic Lung Disease: Review SeriesDocument11 paginiChallenges in Pulmonary Fibrosis ? 3: Cystic Lung Disease: Review SeriesedelinÎncă nu există evaluări
- Bronchiolitis A Practical Approach For The General RadiologistDocument42 paginiBronchiolitis A Practical Approach For The General RadiologistTara NareswariÎncă nu există evaluări
- 2 Complications of Pulmonary TB PDFDocument18 pagini2 Complications of Pulmonary TB PDFGuling SetiawanÎncă nu există evaluări
- Makalah Radiologi TB BE v0.1Document15 paginiMakalah Radiologi TB BE v0.1giovannigyoÎncă nu există evaluări
- Hypothesis", in Which Susceptibility To Infection and Poor Mucociliary Clearance ResultDocument2 paginiHypothesis", in Which Susceptibility To Infection and Poor Mucociliary Clearance ResultAngelique Mae Mortejo SalarzaÎncă nu există evaluări
- Lung Reviewer PathologyDocument9 paginiLung Reviewer PathologyTee VillanuevaÎncă nu există evaluări
- Chest: Recent Advances in Chest MedicineDocument9 paginiChest: Recent Advances in Chest MedicinesmansakobiÎncă nu există evaluări
- Complications of Pulmonary TuberculosisDocument18 paginiComplications of Pulmonary TuberculosisedhobiondiÎncă nu există evaluări
- Materi Kelainan ParuDocument72 paginiMateri Kelainan ParuDebiNingtyasDwiKusumaWardaniÎncă nu există evaluări
- BRONCHIECTASISDocument40 paginiBRONCHIECTASISImmanuel100% (2)
- Etiology: An Irreversible Airway Dilation That Involves The Lung inDocument3 paginiEtiology: An Irreversible Airway Dilation That Involves The Lung inOlli TherineÎncă nu există evaluări
- 15 Signs in Thoracic Imaging.20Document15 pagini15 Signs in Thoracic Imaging.20Don Kihot100% (1)
- Pitfalls of EmphysemaDocument7 paginiPitfalls of EmphysemaWahyu Puspita IrjayantiÎncă nu există evaluări
- Review Article: Expiratory Central Airway Collapse: A Concise ReviewDocument13 paginiReview Article: Expiratory Central Airway Collapse: A Concise ReviewfaberÎncă nu există evaluări
- Case Study: BronchiectasisDocument5 paginiCase Study: BronchiectasisHanna Mae HernandezÎncă nu există evaluări
- PDF To WordDocument17 paginiPDF To Wordanisa ghinaÎncă nu există evaluări
- Evidence-Based Clinical Practice Guidelines Chronic Cough Due To Bronchiectasis: ACCPDocument12 paginiEvidence-Based Clinical Practice Guidelines Chronic Cough Due To Bronchiectasis: ACCPAlizaPinkyÎncă nu există evaluări
- Nader Kamangar, MD, FACP, FCCP FCCM, FAASM, Associate Professor of Clinical Medicine, UniversityDocument7 paginiNader Kamangar, MD, FACP, FCCP FCCM, FAASM, Associate Professor of Clinical Medicine, UniversitynurulfitriantisahÎncă nu există evaluări
- Ultra-Rare Cystic DiseaseDocument17 paginiUltra-Rare Cystic DiseaseRaul DoctoÎncă nu există evaluări
- The Tree in Bud SignDocument2 paginiThe Tree in Bud SignMunish GuleriaÎncă nu există evaluări
- Pictorial Essay: Bronchiectasis: CT CriteriaDocument7 paginiPictorial Essay: Bronchiectasis: CT CriteriadarendraabimayuÎncă nu există evaluări
- Childhood Adult: Bronchiectasis in PulmonaryDocument9 paginiChildhood Adult: Bronchiectasis in PulmonaryphobicmdÎncă nu există evaluări
- Chronic Lower Respiratory Tract InfectionsDocument32 paginiChronic Lower Respiratory Tract Infectionsibnbasheer100% (2)
- Study Guide 2Document16 paginiStudy Guide 2cÎncă nu există evaluări
- BRONKIEKTASIS BaruDocument23 paginiBRONKIEKTASIS BaruRamanda Cahya UmbarraÎncă nu există evaluări
- Parasitic Infections of The Lung: A Guide For The Respiratory PhysicianDocument9 paginiParasitic Infections of The Lung: A Guide For The Respiratory Physicianحسام الدين إسماعيلÎncă nu există evaluări
- J6. Chest X-Ray Manifestations of Pneumonia - Das 2009Document3 paginiJ6. Chest X-Ray Manifestations of Pneumonia - Das 2009Irwan DesriandaÎncă nu există evaluări
- BrochiectasisDocument34 paginiBrochiectasisAndi Wija Indrawan PangerangÎncă nu există evaluări
- DifferentialsDocument7 paginiDifferentialsStewart Garneth Reston RancesÎncă nu există evaluări
- Valeyre2015 Pulmonary SarcoidosisDocument11 paginiValeyre2015 Pulmonary SarcoidosisJohan ArocaÎncă nu există evaluări
- Congenital Lobar Overinflation Congenital Lobar Overinflation, Also KnownDocument3 paginiCongenital Lobar Overinflation Congenital Lobar Overinflation, Also KnownIqbal Muhammad Yaa-BegitulahÎncă nu există evaluări
- A Bronchographic Sign of Chronic: Bronchitis'Document4 paginiA Bronchographic Sign of Chronic: Bronchitis'Ziyan NafisahÎncă nu există evaluări
- B&lung AbscessDocument21 paginiB&lung Abscessnathan asfahaÎncă nu există evaluări
- Patrones Tomograficos Epid PDFDocument28 paginiPatrones Tomograficos Epid PDFYony Morales LeonÎncă nu există evaluări
- Lung Abscess: Presented byDocument36 paginiLung Abscess: Presented byPalanki Gopal100% (1)
- Neu Motor AxDocument6 paginiNeu Motor AxZay SiagÎncă nu există evaluări
- Chronic Obstructive Pulmonary Disease: Iman Galal, MDDocument60 paginiChronic Obstructive Pulmonary Disease: Iman Galal, MDYan Sheng HoÎncă nu există evaluări
- Metod Terapiya Lechebnoe Delo Fakul'tet Inostranyx Studentov-006Document29 paginiMetod Terapiya Lechebnoe Delo Fakul'tet Inostranyx Studentov-006Nishani SatiyaseelanÎncă nu există evaluări
- I Lower Respiratory System PathologyDocument12 paginiI Lower Respiratory System PathologyBaha Al AdaylehÎncă nu există evaluări
- Pathogenic Mechanisms in Asthma and COPDDocument24 paginiPathogenic Mechanisms in Asthma and COPDWilliamRayCassidyÎncă nu există evaluări
- Ajrccm.164.Supplement 2.2106061Document11 paginiAjrccm.164.Supplement 2.2106061Thiago FerreiraÎncă nu există evaluări
- Parasitic Infections of The Lung: A Guide For The Respiratory PhysicianDocument9 paginiParasitic Infections of The Lung: A Guide For The Respiratory PhysicianSuria KumarÎncă nu există evaluări
- (Materi Dr. Heny) PneumoniaDocument86 pagini(Materi Dr. Heny) PneumoniaFathiyah SalsabilaÎncă nu există evaluări
- COP - Cryptogenic Organizing Pneumonia - StatPearls - NCBI BookshelfDocument7 paginiCOP - Cryptogenic Organizing Pneumonia - StatPearls - NCBI Bookshelfrbatjun576Încă nu există evaluări
- Section II - Chest Radiology: Figure 1ADocument35 paginiSection II - Chest Radiology: Figure 1AHaluk Alibazoglu100% (1)
- Medical Mnemonic Sketches : Pulmonary DiseasesDe la EverandMedical Mnemonic Sketches : Pulmonary DiseasesÎncă nu există evaluări
- Anti-Neutrophil Cytoplasmic Antibody (ANCA) Associated VasculitisDe la EverandAnti-Neutrophil Cytoplasmic Antibody (ANCA) Associated VasculitisRenato Alberto SinicoÎncă nu există evaluări
- Salivary Gland Pathology: Diagnosis and ManagementDe la EverandSalivary Gland Pathology: Diagnosis and ManagementÎncă nu există evaluări
- Recurrent Unilateral Pleural Effusion From Constrictive Pericarditis of Unknown Etiology Requiring PericardiectomyDocument3 paginiRecurrent Unilateral Pleural Effusion From Constrictive Pericarditis of Unknown Etiology Requiring PericardiectomyJaya Semara PutraÎncă nu există evaluări
- Aquickreferenceon Respiratoryacidosis: Rebecca A. JohnsonDocument5 paginiAquickreferenceon Respiratoryacidosis: Rebecca A. JohnsonJaya Semara PutraÎncă nu există evaluări
- Lung Hyperinflation in COPD: Applying Physiology To Clinical PracticeDocument12 paginiLung Hyperinflation in COPD: Applying Physiology To Clinical PracticeJaya Semara PutraÎncă nu există evaluări
- PE GuidelinesDocument12 paginiPE GuidelinesJaya Semara PutraÎncă nu există evaluări
- Aztreonam: Antibiotic ClassDocument2 paginiAztreonam: Antibiotic ClassJaya Semara PutraÎncă nu există evaluări
- Post-Tuberculous Bronchiectasis: Indications For Surgical TreatmentDocument2 paginiPost-Tuberculous Bronchiectasis: Indications For Surgical TreatmentJaya Semara PutraÎncă nu există evaluări
- Distress Syndrome (ARDS) : Penggunaan Ventilasi Mekanis Invasif Pada Acute RespiratoryDocument9 paginiDistress Syndrome (ARDS) : Penggunaan Ventilasi Mekanis Invasif Pada Acute Respiratorydoktermuda91Încă nu există evaluări
- AMP-kinase Regulates Food Intake by Responding To Hormonal and Nutrient Signals in The HypothalamusDocument6 paginiAMP-kinase Regulates Food Intake by Responding To Hormonal and Nutrient Signals in The HypothalamusJaya Semara PutraÎncă nu există evaluări
- Zhan El 2004Document7 paginiZhan El 2004Jaya Semara PutraÎncă nu există evaluări
- Respiratory Medicine Case ReportsDocument3 paginiRespiratory Medicine Case ReportsJaya Semara PutraÎncă nu există evaluări
- Streptococcus Suis Serotype 2-Induced Meningoencephalitis: Cerebral Abscesses in A Pig: Atypical Manifestations ofDocument5 paginiStreptococcus Suis Serotype 2-Induced Meningoencephalitis: Cerebral Abscesses in A Pig: Atypical Manifestations ofJaya Semara PutraÎncă nu există evaluări
- 1.BE Airway DefenseDocument11 pagini1.BE Airway DefenseJaya Semara PutraÎncă nu există evaluări
- Immunopathology, Diagnosis, and Management of Hypersensitivity PneumonitisDocument12 paginiImmunopathology, Diagnosis, and Management of Hypersensitivity PneumonitisJaya Semara PutraÎncă nu există evaluări
- Asthma and Pregnancy: Maintenance of Certification Clinical Management SeriesDocument4 paginiAsthma and Pregnancy: Maintenance of Certification Clinical Management SeriesJaya Semara PutraÎncă nu există evaluări
- Balzano Et Al 2001 AllergyDocument8 paginiBalzano Et Al 2001 AllergyJaya Semara PutraÎncă nu există evaluări
- Franco 2017Document12 paginiFranco 2017ReshmaaRajendranÎncă nu există evaluări
- Secreted Giardia Intestinalis Cysteine Proteases Disrupt Intestinal Epithelial Cell JunctionalDocument17 paginiSecreted Giardia Intestinalis Cysteine Proteases Disrupt Intestinal Epithelial Cell JunctionalAura-Nicoleta PopinciucÎncă nu există evaluări
- Cytokine Bioassays PDFDocument383 paginiCytokine Bioassays PDFSubash Suba100% (1)
- Acute & Chronic InflammationDocument102 paginiAcute & Chronic InflammationVenz100% (1)
- Platelet BOOK PDFDocument400 paginiPlatelet BOOK PDFVikashgtm100% (1)
- JCM 08 01283 v2Document44 paginiJCM 08 01283 v2Ambika SuwalÎncă nu există evaluări
- Extending The Host Response' Paradigm From Sepsis To Cardiogenic Shock: Evidence, Limitations and OpportunitiesDocument14 paginiExtending The Host Response' Paradigm From Sepsis To Cardiogenic Shock: Evidence, Limitations and OpportunitiesHelder LezamaÎncă nu există evaluări
- Cellular and Molecular Mechanisms of Asthma and COPDDocument18 paginiCellular and Molecular Mechanisms of Asthma and COPDMihaela BerindeieÎncă nu există evaluări
- Chapter 28 PROMDocument24 paginiChapter 28 PROMTansya PurnaningrumÎncă nu există evaluări
- Ccid 8 239Document10 paginiCcid 8 239Marfatul ArifahÎncă nu există evaluări
- Draft CPG Management of AcneDocument63 paginiDraft CPG Management of AcneAl Mutarabbi100% (1)
- PathDocument31 paginiPathsandhyaÎncă nu există evaluări
- Inflammatory Mediators in The Pathogenesis of Periodontitis: Expert ReviewsDocument22 paginiInflammatory Mediators in The Pathogenesis of Periodontitis: Expert ReviewsPriya MalhotraÎncă nu există evaluări
- 10 Pharmacology - ImmunopharmacologyDocument17 pagini10 Pharmacology - ImmunopharmacologyArnoldBorromeoÎncă nu există evaluări
- Macrofagos Asociados A TumoresDocument9 paginiMacrofagos Asociados A Tumoresjuan ramon zepeda tejedaÎncă nu există evaluări
- Chapter-3 Synthesis, Characterization of Some Novel Chromeno Oxadiazole DerivativesDocument43 paginiChapter-3 Synthesis, Characterization of Some Novel Chromeno Oxadiazole DerivativesMadalina GhitaÎncă nu există evaluări
- Management of COPDDocument30 paginiManagement of COPDDrDishank PatelÎncă nu există evaluări
- Carco C, Young. Incrasing EvidenceDocument24 paginiCarco C, Young. Incrasing Evidenceoliffasalma atthahirohÎncă nu există evaluări
- Acetabular Revision Surgery in MajorDocument223 paginiAcetabular Revision Surgery in MajorWilson A. Quintero100% (2)
- Inflammation, Immunity and Potential Target Therapy of SARS-COV-2 ADocument23 paginiInflammation, Immunity and Potential Target Therapy of SARS-COV-2 AShukr Wesman BlbasÎncă nu există evaluări
- New Concepts On The Pathophysiology of Acute Coronary SyndromeDocument11 paginiNew Concepts On The Pathophysiology of Acute Coronary SyndromeLuis GuerraÎncă nu există evaluări
- Principles of Periodontology Dentino - Et - Al-2013-Periodontology - 2000Document38 paginiPrinciples of Periodontology Dentino - Et - Al-2013-Periodontology - 2000Denisa FloreaÎncă nu există evaluări
- CANINE-Pathophysiology of Organ Failure in Severe Acute Pancreatitis in DogsDocument10 paginiCANINE-Pathophysiology of Organ Failure in Severe Acute Pancreatitis in Dogstaner_soysurenÎncă nu există evaluări
- Pharmacological Therapy of Bronchial Asthma: The Role of BiologicalsDocument12 paginiPharmacological Therapy of Bronchial Asthma: The Role of BiologicalsNia Satila ZariÎncă nu există evaluări
- Seminario 1C Inmunologia General 2023-IDocument16 paginiSeminario 1C Inmunologia General 2023-IPaola Vega GuerraÎncă nu există evaluări
- Pathophysiology and Treatment Syok Septic in PediatricDocument41 paginiPathophysiology and Treatment Syok Septic in PediatricGustomo PanantroÎncă nu există evaluări
- Immune System Dysfunction in The ElderlyDocument16 paginiImmune System Dysfunction in The ElderlyLiya AnjelinaÎncă nu există evaluări
- IF4.02 Molecular Pathogenesis of Klebsiella Pneumoniae PDFDocument36 paginiIF4.02 Molecular Pathogenesis of Klebsiella Pneumoniae PDFdindaÎncă nu există evaluări
- BUUN Immunomodulatory Effects of Massage in Skeletal MuscleDocument133 paginiBUUN Immunomodulatory Effects of Massage in Skeletal Muscledavidescu5costinÎncă nu există evaluări
- New Treatments For Severe Treatment-Resistant Asthma - Lancet Resp Med 2013Document14 paginiNew Treatments For Severe Treatment-Resistant Asthma - Lancet Resp Med 2013Azan Al RasyidÎncă nu există evaluări