S-ar putea să vă placă și
- Polytene and Lampbrush ChromosomesDocument3 paginiPolytene and Lampbrush Chromosomesneeru.bhagatÎncă nu există evaluări
- Chapter 13 Meiosis and Sexual Life CyclesDocument42 paginiChapter 13 Meiosis and Sexual Life Cyclesimma haznaÎncă nu există evaluări
- Biology Tutorial (Set 1)Document18 paginiBiology Tutorial (Set 1)Jin MingÎncă nu există evaluări
- Genetics-Exam 4 1Document5 paginiGenetics-Exam 4 1Farhan Ahmed100% (1)
- Chapter 9 Cell Cycle Chromosome and Mitosis 2018Document42 paginiChapter 9 Cell Cycle Chromosome and Mitosis 2018Nehjoy GerangueÎncă nu există evaluări
- Cell Cycle Cell Division PDFDocument13 paginiCell Cycle Cell Division PDFmuzamil shabir100% (1)
- Genbio1 MitosisDocument11 paginiGenbio1 MitosisJessie Katrisha TayagÎncă nu există evaluări
- Crossing Over: Cc5 AssignmentDocument18 paginiCrossing Over: Cc5 AssignmentAnjishnu Mukhopadhyay BIOTECH SXCBÎncă nu există evaluări
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 paginiCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasÎncă nu există evaluări
- Overview of Cell Cycle by Javali.GDocument15 paginiOverview of Cell Cycle by Javali.GJavali.GÎncă nu există evaluări
- Non DisjunctionDocument59 paginiNon DisjunctionsagaralkimsaimonÎncă nu există evaluări
- BIO 1412 Molecular Biology and Genetics Module - Prof C. Katongo - 2019Document119 paginiBIO 1412 Molecular Biology and Genetics Module - Prof C. Katongo - 2019JOHN MVULA IIÎncă nu există evaluări
- Chapter 4 A Tour of The CellDocument4 paginiChapter 4 A Tour of The Cellmzunl254760% (1)
- Stem Cell Plasticity-Building The Brain of Dreams: by Sally TempleDocument22 paginiStem Cell Plasticity-Building The Brain of Dreams: by Sally TempleGagan KarwarÎncă nu există evaluări
- Ch6a CellDiv Mitosis and Cancer F14sDocument25 paginiCh6a CellDiv Mitosis and Cancer F14sChristopher PhillipsÎncă nu există evaluări
- Cell Cycle Cell Division EditedDocument96 paginiCell Cycle Cell Division Editedmitsuri ʚĭɞÎncă nu există evaluări
- 11th Biology Question BankDocument14 pagini11th Biology Question BankJay senthilÎncă nu există evaluări
- MGS M07 2011 Na CHT A 02Document4 paginiMGS M07 2011 Na CHT A 02rofi modiÎncă nu există evaluări
- ChromosomesDocument4 paginiChromosomesعامر جدونÎncă nu există evaluări
- As Biochemistry QuizDocument7 paginiAs Biochemistry QuizFati QuaynorÎncă nu există evaluări
- The Cell Cycle: Indian Institute of Technology PatnaDocument37 paginiThe Cell Cycle: Indian Institute of Technology PatnaHritik KumarÎncă nu există evaluări
- DNA Topology Supercoiling and Linking PDFDocument5 paginiDNA Topology Supercoiling and Linking PDFmanoj_rkl_07Încă nu există evaluări
- Class-XI Ch-10 Cell Cycle and Cell DivisionDocument12 paginiClass-XI Ch-10 Cell Cycle and Cell DivisionPoonam MeenaÎncă nu există evaluări
- MSC MPlil ZoologyDocument18 paginiMSC MPlil ZoologyYougesh KumarÎncă nu există evaluări
- Mito-Meiosis Test AnswersDocument11 paginiMito-Meiosis Test Answersrosidin_551390Încă nu există evaluări
- Cell DivisionDocument21 paginiCell DivisionOlogunja PaulÎncă nu există evaluări
- MeiosisDocument10 paginiMeiosisNoor Hafezah Mohd MokhtiarÎncă nu există evaluări
- Mammalian Histology AssignmentDocument9 paginiMammalian Histology AssignmentSana Sultana100% (1)
- Regulation of Cell CycleDocument5 paginiRegulation of Cell CyclemohsinÎncă nu există evaluări
- AP Biology Unit 2: CellsDocument67 paginiAP Biology Unit 2: Cellsapi-292966101Încă nu există evaluări
- Nucleotides and Nucleic Acids: Chapter 8 Lehninger 5 EdDocument17 paginiNucleotides and Nucleic Acids: Chapter 8 Lehninger 5 Edarun231187Încă nu există evaluări
- Cell Structures FinalDocument16 paginiCell Structures Finalapi-269480689Încă nu există evaluări
- Cytology McqsDocument8 paginiCytology McqsZiyad AbdallahÎncă nu există evaluări
- Structure of Nuclear Genetic Material - Dna, Rna and Gene: CourseDocument65 paginiStructure of Nuclear Genetic Material - Dna, Rna and Gene: Coursebharath margamÎncă nu există evaluări
- Gene MutationDocument20 paginiGene Mutationdheeresh aggarwalÎncă nu există evaluări
- Meiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyDocument31 paginiMeiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyMariam QaisÎncă nu există evaluări
- The Cell Cycle-TRFDocument8 paginiThe Cell Cycle-TRFmaniiiiiiiiÎncă nu există evaluări
- Mitosis QuestionsDocument10 paginiMitosis QuestionscapremaeÎncă nu există evaluări
- Chapter4 MC QuizDocument19 paginiChapter4 MC Quizsmishra_97Încă nu există evaluări
- Cell Growth and DivisionDocument15 paginiCell Growth and DivisionJohn Lester BerdinÎncă nu există evaluări
- BIOT643 Midterm Exam Summer 2016Document4 paginiBIOT643 Midterm Exam Summer 2016JayÎncă nu există evaluări
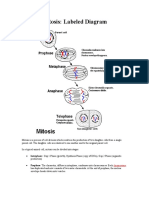
- Mitosis: Labeled Diagram: Interphase: Gap 1 Phase (Growth), Synthesis Phase (Copy of DNA), Gap 2 Phase (OrganelleDocument7 paginiMitosis: Labeled Diagram: Interphase: Gap 1 Phase (Growth), Synthesis Phase (Copy of DNA), Gap 2 Phase (Organelleazzahra adeliaÎncă nu există evaluări
- Karyotyping Virtual Lab PlusDocument3 paginiKaryotyping Virtual Lab PlusVictoria YeeÎncă nu există evaluări
- Chapter 3 - Genetic Control of Cell FunctionDocument7 paginiChapter 3 - Genetic Control of Cell FunctionSADAVEJO100% (1)
- BioSc 231 Exam 3 2003Document9 paginiBioSc 231 Exam 3 2003Quốc ViệtÎncă nu există evaluări
- Cell Division: MitosisDocument38 paginiCell Division: MitosisSam RatnamÎncă nu există evaluări
- Genetics MCQDocument3 paginiGenetics MCQibdpÎncă nu există evaluări
- Linkage, Crossing-Over, & GeneDocument13 paginiLinkage, Crossing-Over, & GeneAbel ClaireÎncă nu există evaluări
- Linkage: Harshraj Subhash Shinde KKW, Cabt, NashikDocument14 paginiLinkage: Harshraj Subhash Shinde KKW, Cabt, Nashiksivaram888Încă nu există evaluări
- Cellular Processes TestDocument4 paginiCellular Processes TestConnorÎncă nu există evaluări
- Module-3 (Theory) Upstream and Down Stream Component of A Fermentation Process PDFDocument6 paginiModule-3 (Theory) Upstream and Down Stream Component of A Fermentation Process PDFAnonymous AgGBWdÎncă nu există evaluări
- Approval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Document12 paginiApproval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Lhely Andi NurlaeliÎncă nu există evaluări
- Bi 341 Chapter 1 The Genetic Code of Genes and Genomes & Introduction - KBDocument76 paginiBi 341 Chapter 1 The Genetic Code of Genes and Genomes & Introduction - KBMATHIXÎncă nu există evaluări
- Bacterial Genetics PDFDocument24 paginiBacterial Genetics PDFJaszmin Sonia KaurÎncă nu există evaluări
- Avatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeDocument2 paginiAvatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeMiguel BernalÎncă nu există evaluări
- NEET Cell Cycle and Cell Divisions Important Questions With Answer Key Free PDFDocument13 paginiNEET Cell Cycle and Cell Divisions Important Questions With Answer Key Free PDFthilaivananÎncă nu există evaluări
- The Pyridine Nucleotide CoenzymesDe la EverandThe Pyridine Nucleotide CoenzymesJohannes EverseÎncă nu există evaluări
- Bacterial ChromosomeDocument2 paginiBacterial Chromosomeharshbharatdesai_783Încă nu există evaluări
- Bacterial ChromosomeDocument2 paginiBacterial Chromosomeharshbharatdesai_783Încă nu există evaluări
- Lalbhai Contractor Stadium MapDocument1 paginăLalbhai Contractor Stadium Mapharshbharatdesai_783Încă nu există evaluări
- Oral Insulin For Diabetes - A Future OptionDocument8 paginiOral Insulin For Diabetes - A Future Optionharshbharatdesai_783Încă nu există evaluări
- 2016, Gene, A Novel Common Large Genomic Deletion and Two New MissenseDocument7 pagini2016, Gene, A Novel Common Large Genomic Deletion and Two New MissenseRomana VulturarÎncă nu există evaluări
- Histopath Transes 1Document9 paginiHistopath Transes 1Nico LokoÎncă nu există evaluări
- Extra Nuclear InheritanceDocument13 paginiExtra Nuclear InheritanceTony BernardÎncă nu există evaluări
- Botânica de AristolochiaDocument13 paginiBotânica de AristolochiaNatã DalarmiÎncă nu există evaluări
- Crop LifeDocument7 paginiCrop Life유가연[학생](생명과학대학 유전생명공학과)Încă nu există evaluări
- Discussion and ConclusionDocument4 paginiDiscussion and ConclusionRisma WerdaningsihÎncă nu există evaluări
- Humoral Immune ResponsesDocument34 paginiHumoral Immune ResponsesvivinÎncă nu există evaluări
- CertificateDocument1 paginăCertificatemrcopy xeroxÎncă nu există evaluări
- GR 12 Life Sciences Participant S Manual Apr May 2018Document126 paginiGR 12 Life Sciences Participant S Manual Apr May 2018lebohangcandy8Încă nu există evaluări
- Nucleic AcidsDocument32 paginiNucleic AcidsAlexander DacumosÎncă nu există evaluări
- Basic Radiobiology (Slides) PDFDocument44 paginiBasic Radiobiology (Slides) PDFSuhey07100% (2)
- Cell Division Activity LabDocument4 paginiCell Division Activity LabSEAN LIBOTÎncă nu există evaluări
- Niper Jee SyllabusDocument6 paginiNiper Jee Syllabuskumar HarshÎncă nu există evaluări
- MCB+102+F12+Lect+ 1+POSTDocument28 paginiMCB+102+F12+Lect+ 1+POSTyoyoma122Încă nu există evaluări
- Notes On Molecular Basis of Inheritance by ExamsRoadDocument8 paginiNotes On Molecular Basis of Inheritance by ExamsRoadVogolus machatteÎncă nu există evaluări
- Clase 1 Biomateriales IBIO 2650Document32 paginiClase 1 Biomateriales IBIO 2650Christian CamiloÎncă nu există evaluări
- GCF 000150735.1 Paracocci BR Pb18 V2 Feature TableDocument1.559 paginiGCF 000150735.1 Paracocci BR Pb18 V2 Feature Tablezero431Încă nu există evaluări
- CARBOHYDRATES With Case StudyDocument1 paginăCARBOHYDRATES With Case StudyGeraldine Roxas CruzÎncă nu există evaluări
- CELL-THE UNIT OF LIFE - LysosomesDocument9 paginiCELL-THE UNIT OF LIFE - LysosomesHridyanshu Singh RoyÎncă nu există evaluări
- Activity 5 WhoDunItDocument3 paginiActivity 5 WhoDunItRuth Danielle GasconÎncă nu există evaluări
- Felix Bronner, Mary C. Farach-Carson, Janet Rubin Bone ResorptionDocument201 paginiFelix Bronner, Mary C. Farach-Carson, Janet Rubin Bone ResorptionCarlos Jibaja Correa100% (1)
- Ultra Structure of ProkaryoteDocument3 paginiUltra Structure of ProkaryoteselvasrijaÎncă nu există evaluări
- Department of Pharmacology Class Routine-April, 2020 (3, 4, 5 & Bds-Semester) Jnims, Porompat ImphalDocument1 paginăDepartment of Pharmacology Class Routine-April, 2020 (3, 4, 5 & Bds-Semester) Jnims, Porompat ImphalNeerajeigya ManoharÎncă nu există evaluări
- Genetics Icar1Document18 paginiGenetics Icar1elanthamizhmaranÎncă nu există evaluări
- trs1003 Annex6Document56 paginitrs1003 Annex6lhthang1990Încă nu există evaluări
- Test Bank For Molecular Biology 5th Edition Robert WeaverDocument36 paginiTest Bank For Molecular Biology 5th Edition Robert Weaverbrokeazarole6rv8pl100% (44)
- DNA MiniprepDocument3 paginiDNA Miniprepifti007Încă nu există evaluări
- Impacts of Gmo'S On Genetically Modified Organisms Produced by Philippine ResearchersDocument3 paginiImpacts of Gmo'S On Genetically Modified Organisms Produced by Philippine ResearchersIrish Cheska EspinosaÎncă nu există evaluări
- Inman Aligner TestimonialsDocument2 paginiInman Aligner TestimonialsInmanAlignerÎncă nu există evaluări
- Sulaiman RT-PCR TestDocument2 paginiSulaiman RT-PCR TestNavid ShadÎncă nu există evaluări