S-ar putea să vă placă și
- Seminario-1 Edema CerebralDocument8 paginiSeminario-1 Edema CerebralANGELA ADRIANA RIVAS SALAZARÎncă nu există evaluări
- Cirugia No Cardiaca en Paciente CardiacoDocument19 paginiCirugia No Cardiaca en Paciente CardiacoGustavo Xavier Ayarza PulidoÎncă nu există evaluări
- Glomerulonefritis Rapidamente Progresiva 2017Document6 paginiGlomerulonefritis Rapidamente Progresiva 2017Sly CribilleroÎncă nu există evaluări
- Documento TraducidoDocument10 paginiDocumento TraducidoYoelys AndreinaÎncă nu există evaluări
- Sindromes PoliuricosDocument33 paginiSindromes PoliuricosMarlene Gonzalez FcoÎncă nu există evaluări
- Causas y síntomas de la hipertensión endocraneanaDocument2 paginiCausas y síntomas de la hipertensión endocraneanaMariSol Pacheco MartinezÎncă nu există evaluări
- Como Escribir El Informe de Un Caso ClinicoDocument3 paginiComo Escribir El Informe de Un Caso ClinicoKaren SalazarÎncă nu există evaluări
- FX InotropicosDocument5 paginiFX InotropicosAnGjBÎncă nu există evaluări
- Glomerulonefritis Rapidamente ProgresivaDocument4 paginiGlomerulonefritis Rapidamente ProgresivaLuciano A. Rurush GuarnizÎncă nu există evaluări
- Edema CerebralDocument7 paginiEdema CerebralVictor OrozcoÎncă nu există evaluări
- SLEDAI - Indicadores de Actividad LESDocument9 paginiSLEDAI - Indicadores de Actividad LESpedrixon123Încă nu există evaluări
- Coeficiente de Correlacion LinealDocument5 paginiCoeficiente de Correlacion LinealRosie Paz100% (1)
- Anemia PerniciosaDocument13 paginiAnemia PerniciosaAQUACOSTA SASÎncă nu există evaluări
- Glomerulonefritis Rápidamente ProgresivaDocument9 paginiGlomerulonefritis Rápidamente ProgresivaYamil FernandezÎncă nu există evaluări
- Cuadro Células y Organos Del Sistema InmuneDocument12 paginiCuadro Células y Organos Del Sistema InmuneDraco10110101Încă nu există evaluări
- Inflamación y ReparaciónDocument120 paginiInflamación y ReparaciónErick Antonio Castillo GurdianÎncă nu există evaluări
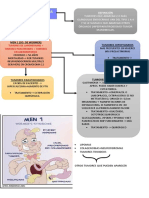
- Neoplasia Endocrina MultipleDocument2 paginiNeoplasia Endocrina MultipleClaudia VargasÎncă nu există evaluări
- Anestesico Endovenoso DaltonDocument90 paginiAnestesico Endovenoso DaltonSheyla Ascencio OkonÎncă nu există evaluări
- Corazón PDF 1Document145 paginiCorazón PDF 1Omar Leonardo Guerrero SuarezÎncă nu există evaluări
- Enfermedad pulmonar intersticial difusa: fisiopatología y diagnósticoDocument54 paginiEnfermedad pulmonar intersticial difusa: fisiopatología y diagnósticoLuzmila Guillinta HernándezÎncă nu există evaluări
- Absceso CerebralDocument7 paginiAbsceso Cerebralwww.pacourgencias.blogspot.com100% (1)
- Esclerodermia Revision 2020Document4 paginiEsclerodermia Revision 2020Jhon HAÎncă nu există evaluări
- Hemorragias de Vías Digestivas Altas y BajasDocument13 paginiHemorragias de Vías Digestivas Altas y BajasSergio ArbelaezÎncă nu există evaluări
- Edema CerebalDocument7 paginiEdema CerebalValentina CárdenasÎncă nu există evaluări
- Cardiopatias Congenitas CianoticasDocument35 paginiCardiopatias Congenitas CianoticasAngélica Huerta100% (1)
- Anemia AplásicaDocument5 paginiAnemia AplásicaNEYDA QUISPE CARHUANIÎncă nu există evaluări
- Hemoptisis - Lucas RomanDocument9 paginiHemoptisis - Lucas RomanLucas RománÎncă nu există evaluări
- Falla Orgánica MúltipleDocument8 paginiFalla Orgánica MúltipleGiordan SanchezÎncă nu există evaluări
- Fagocitosis Universidad Nacional Federico VillarrealDocument4 paginiFagocitosis Universidad Nacional Federico VillarrealAngel Ramiro Soplin AlmeidaÎncă nu există evaluări
- Anatomia Pulmonar NormalDocument33 paginiAnatomia Pulmonar Normalpaor84100% (1)
- Como Hace Un Caso ClinicoDocument2 paginiComo Hace Un Caso ClinicoManuel Alejandro Romero PerezÎncă nu există evaluări
- Anemias HemoliticasDocument37 paginiAnemias HemoliticasHarrinson Requejo CobaÎncă nu există evaluări
- Anemias Hemolíticas Por Alteraciónes de Enzimas EritrocitariasDocument13 paginiAnemias Hemolíticas Por Alteraciónes de Enzimas EritrocitariasGaby ReyesÎncă nu există evaluări
- NeumocistosisDocument13 paginiNeumocistosisMichel HernándezÎncă nu există evaluări
- Síndromes poliúricos: causas y abordajeDocument28 paginiSíndromes poliúricos: causas y abordajeCecii Sejas Guzmán100% (1)
- Guía actualizada sobre EPOCDocument11 paginiGuía actualizada sobre EPOCNatalia Valenzuela ValenzuelaÎncă nu există evaluări
- Lesiones Del Espacio Aéreo - Alejandra MarquezDocument8 paginiLesiones Del Espacio Aéreo - Alejandra MarquezAlejandra MárquezÎncă nu există evaluări
- Perfil Cardiovascular en Pacientes en ShockDocument7 paginiPerfil Cardiovascular en Pacientes en Shockedson martin giron vazquezÎncă nu există evaluări
- 4 Causas Fisiopatologicas Del Fallo Respiratorio AgudoDocument9 pagini4 Causas Fisiopatologicas Del Fallo Respiratorio Agudoapi-3697492Încă nu există evaluări
- ALTERACIONESDocument18 paginiALTERACIONESChipiRosalesÎncă nu există evaluări
- Trabajo Integrador Final - Sistema RespiratorioDocument16 paginiTrabajo Integrador Final - Sistema RespiratorioGabriel Carp PalazzoloÎncă nu există evaluări
- Defensa frente a cambios de pHDocument58 paginiDefensa frente a cambios de pHOmar Alexei Monge de YtaÎncă nu există evaluări
- Clase Sindrome Mieloproliferativos Parte 1Document48 paginiClase Sindrome Mieloproliferativos Parte 1Tell Me DaryÎncă nu există evaluări
- Insuficiencia Cardiaca UrgenciasDocument49 paginiInsuficiencia Cardiaca UrgenciasJose angel Rojas martinez100% (1)
- Gasometría arterial y venosa: Diferencias prácticasDocument4 paginiGasometría arterial y venosa: Diferencias prácticasJovanni Samuel Vazquez IbañezÎncă nu există evaluări
- Síndrome de Distrés Respiratorio Agudo (Sdra)Document46 paginiSíndrome de Distrés Respiratorio Agudo (Sdra)Silvia Huañapaco YujraÎncă nu există evaluări
- Hipertensión Arterial Sistémica Curso Integral de AcreditaciónDocument215 paginiHipertensión Arterial Sistémica Curso Integral de AcreditaciónFlamengod100% (1)
- Alergia A MedicamentosDocument38 paginiAlergia A MedicamentosAntonella Castro CabreraÎncă nu există evaluări
- EspirometríaDocument17 paginiEspirometríaMarianela Vásquez Araya100% (1)
- Capitulo 2 Electrofisiologia CelularDocument6 paginiCapitulo 2 Electrofisiologia CelularÁlvaro RuedaÎncă nu există evaluări
- Ciclo cardíaco: resumen de sus fases y eventosDocument3 paginiCiclo cardíaco: resumen de sus fases y eventosCarlos Alexander A A100% (1)
- Anemias Hemolíticas Por Alteraciones de Enzimas EritrocitariasDocument43 paginiAnemias Hemolíticas Por Alteraciones de Enzimas EritrocitariasAbyYadoHuertaÎncă nu există evaluări
- Neumonía Adquirida en La ComunidadDocument33 paginiNeumonía Adquirida en La ComunidadGabriela Rubio WestermeierÎncă nu există evaluări
- Neumopatías Intersticiales DifusasDocument33 paginiNeumopatías Intersticiales DifusasCarlos M Garcia LaraÎncă nu există evaluări
- Meningitis Aguda en NeonatosDocument3 paginiMeningitis Aguda en NeonatosMireya RamirezÎncă nu există evaluări
- Linfoma no Hodgkin en niños: causas, síntomas y tratamientoDocument12 paginiLinfoma no Hodgkin en niños: causas, síntomas y tratamientowilmer valladaresÎncă nu există evaluări
- Manual para entender y tratar el asma: Consejos para mejorar la calidad de vidaDe la EverandManual para entender y tratar el asma: Consejos para mejorar la calidad de vidaÎncă nu există evaluări
- Quimica Uabp 3 EtanolDocument7 paginiQuimica Uabp 3 EtanolOsvaldo HerraezÎncă nu există evaluări
- Estética de UñasDocument4 paginiEstética de UñasAladino AbbasÎncă nu există evaluări
- Keratina para El CabelloDocument5 paginiKeratina para El CabelloAladino AbbasÎncă nu există evaluări
- El Mejor Monómero para Uñas AcrílicasDocument5 paginiEl Mejor Monómero para Uñas AcrílicasAladino AbbasÎncă nu există evaluări
- Bombas Efervescentes para PiesDocument5 paginiBombas Efervescentes para PiesAladino AbbasÎncă nu există evaluări
- Uñas AcrílicasDocument8 paginiUñas AcrílicasAladino AbbasÎncă nu există evaluări
- La AlquimiaDocument7 paginiLa AlquimiaAladino AbbasÎncă nu există evaluări
- Padrastro (Uña)Document1 paginăPadrastro (Uña)Aladino AbbasÎncă nu există evaluări
- Estructura de La Uña Natural y Anatomía de La UñaDocument8 paginiEstructura de La Uña Natural y Anatomía de La UñaAladino AbbasÎncă nu există evaluări
- ParoniquiaDocument3 paginiParoniquiaAladino AbbasÎncă nu există evaluări
- Átomo 1Document14 paginiÁtomo 1Aladino AbbasÎncă nu există evaluări
- AlquimiaDocument22 paginiAlquimiaAladino AbbasÎncă nu există evaluări
- AlquimiaDocument22 paginiAlquimiaAladino AbbasÎncă nu există evaluări
- AlquimiaDocument22 paginiAlquimiaAladino AbbasÎncă nu există evaluări
- Orbital AtómicoDocument5 paginiOrbital AtómicoAladino AbbasÎncă nu există evaluări
- Protón 3Document5 paginiProtón 3Aladino AbbasÎncă nu există evaluări
- Neutrón 4Document4 paginiNeutrón 4Aladino AbbasÎncă nu există evaluări
- Electrón 2Document30 paginiElectrón 2Aladino AbbasÎncă nu există evaluări
- Galactose MiaDocument10 paginiGalactose MiaAladino AbbasÎncă nu există evaluări
- HematuriaDocument8 paginiHematuriaAladino Abbas100% (1)
- Biología WikiDocument15 paginiBiología WikiAladino AbbasÎncă nu există evaluări
- Biofísica WikiDocument4 paginiBiofísica WikiAladino AbbasÎncă nu există evaluări
- Análisis de Sedimentos en La OrinaDocument5 paginiAnálisis de Sedimentos en La OrinaAladino AbbasÎncă nu există evaluări
- Gregor MendelDocument8 paginiGregor MendelAladino AbbasÎncă nu există evaluări
- 09 El BigbangDocument196 pagini09 El BigbangRey LagartoÎncă nu există evaluări
- Lactasa PDFDocument6 paginiLactasa PDFAladino AbbasÎncă nu există evaluări
- Ga LactosaDocument2 paginiGa LactosaAladino AbbasÎncă nu există evaluări
- 09 El BigbangDocument196 pagini09 El BigbangRey LagartoÎncă nu există evaluări
- Lactasa PDFDocument6 paginiLactasa PDFAladino AbbasÎncă nu există evaluări
- Lactasa PDFDocument6 paginiLactasa PDFAladino AbbasÎncă nu există evaluări
- LactosaDocument4 paginiLactosaAladino AbbasÎncă nu există evaluări
- Ensayo 4 Medio Mencion QuimicaDocument20 paginiEnsayo 4 Medio Mencion QuimicaMaría TemucoÎncă nu există evaluări
- Wuolah Free Apuntes BioquimicaDocument118 paginiWuolah Free Apuntes Bioquimicajavier saizÎncă nu există evaluări
- Glosario FarmacoDocument41 paginiGlosario FarmacoLuz CalvaÎncă nu există evaluări
- P4 Dulzor de AzucaresDocument14 paginiP4 Dulzor de AzucaresAnel MamaniÎncă nu există evaluări
- El ArrozDocument9 paginiEl ArrozSilverÎncă nu există evaluări
- Glúcidos: Monosacáridos y Disacáridos de Interés BiológicoDocument29 paginiGlúcidos: Monosacáridos y Disacáridos de Interés BiológicoPatriciaÎncă nu există evaluări
- BIOCÁPSULA CarbohidratosDocument5 paginiBIOCÁPSULA CarbohidratosMaria Milagro Arteaga EscalonaÎncă nu există evaluări
- Crucigrama Alimentación y Vida Primer ParcialDocument2 paginiCrucigrama Alimentación y Vida Primer ParcialNicole ZehennyÎncă nu există evaluări
- Bases Alimentarias y NutriciónDocument19 paginiBases Alimentarias y NutriciónJohanna EscobarÎncă nu există evaluări
- COCA Y DIETA CETOGENICA - Altern - Kunihiro Seki PH.DDocument142 paginiCOCA Y DIETA CETOGENICA - Altern - Kunihiro Seki PH.DConstanza Alvarez100% (4)
- Silabo Bioquimica 2021Document7 paginiSilabo Bioquimica 2021San KanÎncă nu există evaluări
- HabasDocument117 paginiHabasAna Rodriguez Rodriguez100% (1)
- Gomas Y Pectinas.Document19 paginiGomas Y Pectinas.Christopher Daniel Flores HernándezÎncă nu există evaluări
- La Importancia Dela Nutrición para La Actividad Física en Pacientes Críticamente EnfermosDocument10 paginiLa Importancia Dela Nutrición para La Actividad Física en Pacientes Críticamente EnfermosSinahí MaroteÎncă nu există evaluări
- Polisacáridos: macromoléculas formadas por monosacáridosDocument55 paginiPolisacáridos: macromoléculas formadas por monosacáridosAna LeonÎncă nu există evaluări
- Calculo FormatoDocument3 paginiCalculo FormatoFierro Villa Valeria GuadalupeÎncă nu există evaluări
- Nutrición en Atletas de Alto RendimientoDocument3 paginiNutrición en Atletas de Alto RendimientoLuis RoblesÎncă nu există evaluări
- Sprinters CubaDocument11 paginiSprinters CubaJorge Airy MercadoÎncă nu există evaluări
- Librosusc5, Leches Azucaradas-65-92Document28 paginiLibrosusc5, Leches Azucaradas-65-92johaÎncă nu există evaluări
- Quimica de Las MacromoleculasDocument119 paginiQuimica de Las MacromoleculasCarlos Urgiles RollandÎncă nu există evaluări
- Reconocimiento de Biomoleculas FINALDocument19 paginiReconocimiento de Biomoleculas FINALPepe Rojas Yachas100% (1)
- CuestionarioDocument7 paginiCuestionarioSayuri Elena Petro MaceaÎncă nu există evaluări
- Informe #05Document10 paginiInforme #05Liliana AlvaradoÎncă nu există evaluări
- Informe Azucares ReductoresDocument6 paginiInforme Azucares ReductoresDayana Romero MirandaÎncă nu există evaluări
- PRÁCTICA DE CARBOHIDRATOS: RECONOCIMIENTO DE AZÚCARESDocument15 paginiPRÁCTICA DE CARBOHIDRATOS: RECONOCIMIENTO DE AZÚCARESJuan C ZavaletaÎncă nu există evaluări
- Guia Ciencias Grado 7° 2° Periodo 2020Document21 paginiGuia Ciencias Grado 7° 2° Periodo 2020daiidiÎncă nu există evaluări
- Diapositivas Organica-CarbohidratosDocument36 paginiDiapositivas Organica-CarbohidratosmaferChevezÎncă nu există evaluări
- Guia de Laboratorio - Reconocimiento de CarbohidratosDocument3 paginiGuia de Laboratorio - Reconocimiento de CarbohidratosDeius RuizÎncă nu există evaluări
- Grados Brix y Prueba de FehlingDocument4 paginiGrados Brix y Prueba de FehlingRicardo Franco ArboledaÎncă nu există evaluări
- Anexo 3 Quimica (Recuperado Automáticamente)Document23 paginiAnexo 3 Quimica (Recuperado Automáticamente)Kely Marroquin Rozo100% (1)