S-ar putea să vă placă și
- Paso 5 Propuesta de Manejo Biologico en Mora - Grupo 13Document14 paginiPaso 5 Propuesta de Manejo Biologico en Mora - Grupo 13Julian Tovar MedinaÎncă nu există evaluări
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularDe la EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularÎncă nu există evaluări
- Qué Es Una CélulaDocument18 paginiQué Es Una CélulaJohn EderÎncă nu există evaluări
- Cuadernillo de Biologia 1 Bloque Iii - ZambranoDocument23 paginiCuadernillo de Biologia 1 Bloque Iii - ZambranoMarcos Zambrano AlegriaÎncă nu există evaluări
- Atencio N A La Diversidad 4º ESO Biologi A y Geologi A PDFDocument64 paginiAtencio N A La Diversidad 4º ESO Biologi A y Geologi A PDFJose Maria Benito Garcia100% (1)
- Prueba de Diagnostico Segundos Medios BioDocument1 paginăPrueba de Diagnostico Segundos Medios BioJavier Ignacio Acevedo MorenoÎncă nu există evaluări
- Unidad1 ReplicacionygenesDocument41 paginiUnidad1 ReplicacionygenesRockdrigo ArellanoÎncă nu există evaluări
- Guía TP 5Document16 paginiGuía TP 5Dani Morales DellatorreÎncă nu există evaluări
- 6551-Libro 1 Parte Ii-Bio Molecular y Celular-Biología Mención 2018 - 7% PDFDocument153 pagini6551-Libro 1 Parte Ii-Bio Molecular y Celular-Biología Mención 2018 - 7% PDFCAMILA HUINCAHUEÎncă nu există evaluări
- Tarea 1 BiologiaDocument12 paginiTarea 1 BiologiaAngie Lizeth Oviedo BautistaÎncă nu există evaluări
- Guía Biología C - Octubre 2016 - Versión ColorDocument139 paginiGuía Biología C - Octubre 2016 - Versión ColorBel Minda CostaÎncă nu există evaluări
- Guía Biología C - Octubre 2016 - Versión Color PDFDocument139 paginiGuía Biología C - Octubre 2016 - Versión Color PDFAlvaro Diego CastilloÎncă nu există evaluări
- MATERIALDocument24 paginiMATERIALJVCLUISÎncă nu există evaluări
- Generalidades de La CélulaDocument86 paginiGeneralidades de La CélulaIsabella Villanueva GallegoÎncă nu există evaluări
- Cta1 U1-Sesion3Document8 paginiCta1 U1-Sesion3Kristhen Talia Zuiicide LinceÎncă nu există evaluări
- Tercer Informe de BiologiaDocument15 paginiTercer Informe de BiologiaMichelle BustosÎncă nu există evaluări
- Cta1 U1-Sesion3Document7 paginiCta1 U1-Sesion3SELENI CHOQUEPUMA TACOÎncă nu există evaluări
- Unidad3.Variabilidadgenetica 131216Document37 paginiUnidad3.Variabilidadgenetica 131216enriqueILAPÎncă nu există evaluări
- Variabilidad GenéticaDocument37 paginiVariabilidad GenéticaPedro RijasÎncă nu există evaluări
- De Acuerdo A Esto Vamos A Definir A Las Células ComoDocument4 paginiDe Acuerdo A Esto Vamos A Definir A Las Células ComoPatricio CarrÎncă nu există evaluări
- Biología Unidad - 1Document19 paginiBiología Unidad - 1Leider Hernandez HZÎncă nu există evaluări
- Bíología Molecular y GenéticaDocument22 paginiBíología Molecular y GenéticaMaria Jose Rodriguez GaonaÎncă nu există evaluări
- Capitulo 9Document46 paginiCapitulo 9Leonela AmayaÎncă nu există evaluări
- 4 PDFDocument5 pagini4 PDFGuisellaÎncă nu există evaluări
- La Teoría Celular BiologiaDocument11 paginiLa Teoría Celular BiologiaIvan QuiñonesÎncă nu există evaluări
- Tarea 2 Mapas ConceptualesDocument14 paginiTarea 2 Mapas ConceptualesMelany VivancoÎncă nu există evaluări
- Cta2 U1 Sesión 02Document7 paginiCta2 U1 Sesión 02Daysi CarmenÎncă nu există evaluări
- Actividad Silabo 3 BiologiaDocument8 paginiActividad Silabo 3 BiologiaDaniel Felipe Olivera GutierrezÎncă nu există evaluări
- Ejercicios2 4 PD BioDocument2 paginiEjercicios2 4 PD BiokarinÎncă nu există evaluări
- 1 TALLER. Generalidades de La Célula y Su EstructuraDocument24 pagini1 TALLER. Generalidades de La Célula y Su EstructuraCamilo andres mejia vasquezÎncă nu există evaluări
- Microbiología - Guía Pedagógica2Document6 paginiMicrobiología - Guía Pedagógica2Bautista CofréÎncă nu există evaluări
- Syllabus Bi-183 - Biologia Celular - 2022-1Document4 paginiSyllabus Bi-183 - Biologia Celular - 2022-1YEFERSON WILMER QUISPE CERDAÎncă nu există evaluări
- PraticaDocument13 paginiPraticaYazz NLopÎncă nu există evaluări
- GuiaDocument20 paginiGuiabiologorogerÎncă nu există evaluări
- Apunte ADN - Época 2 - 2024Document6 paginiApunte ADN - Época 2 - 2024galvanmaximo315Încă nu există evaluări
- CIENCIA Y TECNOLOGÍA - Miércoles 05 de Abril - V CICLO - 2023Document10 paginiCIENCIA Y TECNOLOGÍA - Miércoles 05 de Abril - V CICLO - 2023Kiara Candelario huaytallaÎncă nu există evaluări
- Arti 1. 2023 TP N1. Biomoléculas y CélulaDocument4 paginiArti 1. 2023 TP N1. Biomoléculas y CélulaHoracio LlopÎncă nu există evaluări
- PraticaDocument13 paginiPraticaYazz NLopÎncă nu există evaluări
- GUIA 2 BIOLOGIA 8o. 2020Document5 paginiGUIA 2 BIOLOGIA 8o. 2020Li NaÎncă nu există evaluări
- N° 1Document8 paginiN° 1irina ortegaÎncă nu există evaluări
- Secuencia Didáctica CélulaDocument7 paginiSecuencia Didáctica CélulaCarina Sanchis100% (1)
- ANGEL P.3.Cuadernillo - Biología-1 - 22-23ADocument5 paginiANGEL P.3.Cuadernillo - Biología-1 - 22-23ARenata Herrera GalvanÎncă nu există evaluări
- Unidad Educativa Liceo Cristiano de Guayaquil: Taller 1 de Biologia: TemaDocument12 paginiUnidad Educativa Liceo Cristiano de Guayaquil: Taller 1 de Biologia: TemaDeisy JTigreÎncă nu există evaluări
- Microcurriculo de La Celula Grado SextoDocument13 paginiMicrocurriculo de La Celula Grado Sextoibethrengifo8Încă nu există evaluări
- Tópicos Selectos de Biología I. Segundo ParcialDocument55 paginiTópicos Selectos de Biología I. Segundo ParcialJOSE GUADALUPEÎncă nu există evaluări
- Biologia MolescularDocument8 paginiBiologia MolescularyuriÎncă nu există evaluări
- Las Células CLASE 4 VIRTUALDocument6 paginiLas Células CLASE 4 VIRTUALlili pederneraÎncă nu există evaluări
- Guía de Trabajo CelularDocument9 paginiGuía de Trabajo Celularjesus clementtÎncă nu există evaluări
- Guía de TP 1 2021Document8 paginiGuía de TP 1 2021Florencia HautevilleÎncă nu există evaluări
- TP1 ParcialDocument8 paginiTP1 ParcialJosé Neto50% (2)
- PLAN CuartoDocument161 paginiPLAN Cuartonatalia andrea uribeÎncă nu există evaluări
- Lectura1 2mediopdf 140409155316 Phpapp02Document11 paginiLectura1 2mediopdf 140409155316 Phpapp02Roxana Alvarez DuranÎncă nu există evaluări
- Lab 9. División Celular IDocument10 paginiLab 9. División Celular IChristian Preciado RetoÎncă nu există evaluări
- Actividad de Sociabilización - Seres Vivos - CWZ - 2017Document5 paginiActividad de Sociabilización - Seres Vivos - CWZ - 2017paula arayaÎncă nu există evaluări
- Sesion 2 Celulas Procariotas..Document9 paginiSesion 2 Celulas Procariotas..Franklin Muñoz HilaresÎncă nu există evaluări
- Guía de TP 1 2022Document8 paginiGuía de TP 1 2022josito7u7Încă nu există evaluări
- 1.2 Panorámica Gral. Célula CuestionarioDocument6 pagini1.2 Panorámica Gral. Célula CuestionarioRiver RiverÎncă nu există evaluări
- Guía de TP 1 2022Document8 paginiGuía de TP 1 2022estefany milagrosÎncă nu există evaluări
- P6 Semana Celula EucariotaDocument2 paginiP6 Semana Celula EucariotaGisela Barrios PorrasÎncă nu există evaluări
- Guia de Estudios y Trabajos Prácticos LibresDocument16 paginiGuia de Estudios y Trabajos Prácticos LibresMarta Luciana RoldanÎncă nu există evaluări
- Bibp U1 Ea HuocDocument5 paginiBibp U1 Ea Huochugo ortegaÎncă nu există evaluări
- Bou2 U1 A2 HuocDocument3 paginiBou2 U1 A2 Huochugo ortegaÎncă nu există evaluări
- Bou2 U1 Atr HuocDocument2 paginiBou2 U1 Atr Huochugo ortegaÎncă nu există evaluări
- Bibp1 U1 Ea HuocDocument4 paginiBibp1 U1 Ea Huochugo ortegaÎncă nu există evaluări
- BIBP1 U1 ContenidoDocument102 paginiBIBP1 U1 Contenidohugo ortegaÎncă nu există evaluări
- Bibp1 U1 Atr HuocDocument2 paginiBibp1 U1 Atr Huochugo ortegaÎncă nu există evaluări
- Bibp1 U1 A2 HuocDocument5 paginiBibp1 U1 A2 Huochugo ortegaÎncă nu există evaluări
- Qué Es Un Lecho PorosoDocument1 paginăQué Es Un Lecho Porosohugo ortegaÎncă nu există evaluări
- Bibp1 U2 A1 HuocDocument6 paginiBibp1 U2 A1 Huochugo ortegaÎncă nu există evaluări
- Tesoro 20Document16 paginiTesoro 20hugo ortegaÎncă nu există evaluări
- Bfde U3 Ea3 OchuDocument3 paginiBfde U3 Ea3 Ochuhugo ortega100% (2)
- Endotermos vs. EctodermosDocument8 paginiEndotermos vs. EctodermosKarla Dennise Guerra CruzÎncă nu există evaluări
- Tema 20: Defensas Del Organismo Frente A InfeccionesDocument9 paginiTema 20: Defensas Del Organismo Frente A Infeccionesnana31Încă nu există evaluări
- Una Guía para La Aplicación: Fumigantes Del Suelo Agrícola TeloneDocument60 paginiUna Guía para La Aplicación: Fumigantes Del Suelo Agrícola TeloneWilder Vergara CastañoÎncă nu există evaluări
- Flor de CafeDocument11 paginiFlor de Cafejohann roldan100% (1)
- Primera Guerra Mundial Trabajo Primero MedioDocument6 paginiPrimera Guerra Mundial Trabajo Primero MedioTomás ReyesÎncă nu există evaluări
- CUESTIONARIO MicrobiologíaDocument2 paginiCUESTIONARIO MicrobiologíaJesús Antonio Reyes MejiaÎncă nu există evaluări
- Produccion Acido CitricoDocument5 paginiProduccion Acido CitricobobwayneÎncă nu există evaluări
- 5 Rabdomiosarcomas y No RabdoDocument5 pagini5 Rabdomiosarcomas y No RabdoClaudia MadrigalÎncă nu există evaluări
- Proyecto Edwin Perdomo ParisDocument41 paginiProyecto Edwin Perdomo Parisedwin 012Încă nu există evaluări
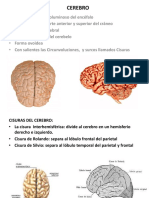
- CerebroDocument10 paginiCerebroandreaB100% (1)
- Actividad 1 (13) (Reparado)Document14 paginiActividad 1 (13) (Reparado)Milena Grisales LòpezÎncă nu există evaluări
- Taller de Interfaz ExcelDocument5 paginiTaller de Interfaz ExcelAnderson MirandaÎncă nu există evaluări
- Nuerodinamia ResumenDocument8 paginiNuerodinamia Resumendomehc17022002Încă nu există evaluări
- Colombia Magia SalvajeDocument4 paginiColombia Magia SalvajeJean Carlos TolozaÎncă nu există evaluări
- ZZZ (T) - Medicina Tradicional y Uso de Plantas Medicinales en Los Cantones Antonio Ante y Cotacachi, (2015) PDFDocument91 paginiZZZ (T) - Medicina Tradicional y Uso de Plantas Medicinales en Los Cantones Antonio Ante y Cotacachi, (2015) PDFJuanManuelAmaroLuis100% (1)
- Primeros AuxiliosDocument33 paginiPrimeros AuxiliosEstefania HolguinÎncă nu există evaluări
- HematologiaDocument56 paginiHematologiaArnoldo FelixÎncă nu există evaluări
- Sistemas de ClasificaciónDocument10 paginiSistemas de ClasificaciónabigailÎncă nu există evaluări
- Porcicultura Autor Myriam Gisella Cruz ChuquitarcoDocument57 paginiPorcicultura Autor Myriam Gisella Cruz ChuquitarcoFELIPE RONALDO ROSADO SALTOSÎncă nu există evaluări
- Derecho AmbientalDocument13 paginiDerecho AmbientalRaquel Alejandra Campos UmañaÎncă nu există evaluări
- Sesion 2 Litosfera y Ecos TerrestreDocument78 paginiSesion 2 Litosfera y Ecos Terrestrefernando felipe reyes castilloÎncă nu există evaluări
- Concord ft-058 PDFDocument5 paginiConcord ft-058 PDFRafael BonavidesÎncă nu există evaluări
- Examen BioiiDocument7 paginiExamen BioiiBiol Itzayara Novoa SerranoÎncă nu există evaluări
- Practica de Laboratorio 03 Hoja MODIFICADADocument12 paginiPractica de Laboratorio 03 Hoja MODIFICADAEmelyn SaldivarÎncă nu există evaluări
- 7 Complejidad y Organización de Los SistemasDocument8 pagini7 Complejidad y Organización de Los SistemasRoddy VicenteÎncă nu există evaluări
- Evaluacion FisioterapeuticaDocument17 paginiEvaluacion Fisioterapeuticabrian albert aaron nuñez melgar rojasÎncă nu există evaluări
- Meiosis y MitosisDocument12 paginiMeiosis y MitosisSergio Rios ContrerasÎncă nu există evaluări
- Guía Práctica Morfología MicroscopicaDocument10 paginiGuía Práctica Morfología MicroscopicaMichelle Oriana Angarita GonzalezÎncă nu există evaluări
- APUNTES CLASES - HistologiaDocument7 paginiAPUNTES CLASES - HistologiaValeria ArbelaezÎncă nu există evaluări