S-ar putea să vă placă și
- Skullbasefracturesand Theircomplications: Kristen L. Baugnon,, Patricia A. HudginsDocument27 paginiSkullbasefracturesand Theircomplications: Kristen L. Baugnon,, Patricia A. Hudginshenandwitafadilla28Încă nu există evaluări
- Growth and Development of Cranial Base: SeminarDocument34 paginiGrowth and Development of Cranial Base: SeminarMNSÎncă nu există evaluări
- The Anterior and Middle Cranial BaseDocument30 paginiThe Anterior and Middle Cranial Basejuan valencia sotoÎncă nu există evaluări
- Head and Neck Cheat Sheets PDFDocument8 paginiHead and Neck Cheat Sheets PDFJM LiensÎncă nu există evaluări
- Chapter 139 Anatomy of The Skull Base Temporal Bone External Ear and Middle Ear Larry G Duckert - CompressDocument12 paginiChapter 139 Anatomy of The Skull Base Temporal Bone External Ear and Middle Ear Larry G Duckert - CompressWudieÎncă nu există evaluări
- Essay Solution On Space in The SkullDocument21 paginiEssay Solution On Space in The SkullEmmanuel IshiomaÎncă nu există evaluări
- Cosmetic Otoplasty 2018 DBDocument11 paginiCosmetic Otoplasty 2018 DBcirugia plastica uisÎncă nu există evaluări
- Bones of The Skull Author Department of Oral and Maxillofacial SurgeryDocument17 paginiBones of The Skull Author Department of Oral and Maxillofacial Surgeryمدونة الأحترافÎncă nu există evaluări
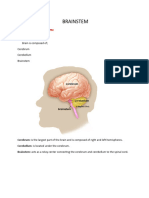
- BRAINSTEMDocument17 paginiBRAINSTEMaroosadilbar27Încă nu există evaluări
- Ns1 Assignment 2 Meninges Group 1Document4 paginiNs1 Assignment 2 Meninges Group 1Roselle AbrazaldoÎncă nu există evaluări
- GRDA Intro Head&NeckDocument18 paginiGRDA Intro Head&NeckKingÎncă nu există evaluări
- Osta Lecture 4 Notes Online ENGLISH Base of Skull and BrainDocument36 paginiOsta Lecture 4 Notes Online ENGLISH Base of Skull and BrainslyfoxkittyÎncă nu există evaluări
- Craneo Sobotta PDFDocument111 paginiCraneo Sobotta PDFMaria Teresa AlvaradoÎncă nu există evaluări
- Anterior and Posterior Petrosectomy Seminar 13 Jan 2021 DR Abhinav 2Document76 paginiAnterior and Posterior Petrosectomy Seminar 13 Jan 2021 DR Abhinav 2Tamajyoti GhoshÎncă nu există evaluări
- Mgutknecht,+sdj Article 618Document13 paginiMgutknecht,+sdj Article 618Emanuela MironÎncă nu există evaluări
- Exterior and Middle EarDocument26 paginiExterior and Middle EarAndra BauerÎncă nu există evaluări
- Aprimaion Angulo PontocerebelosoDocument64 paginiAprimaion Angulo PontocerebelosoLuis Carlos Avellaneda CurchoÎncă nu există evaluări
- Block 3 Review Slides-PART 01 (2 Files Merged)Document222 paginiBlock 3 Review Slides-PART 01 (2 Files Merged)spitzmark2030Încă nu există evaluări
- Anatomyofthe Auditory SystemDocument6 paginiAnatomyofthe Auditory System32. Nguyễn Tiến Thái SơnÎncă nu există evaluări
- Anatomy 1Document1 paginăAnatomy 1Brian RamosÎncă nu există evaluări
- Cranial BaseDocument25 paginiCranial BasesealovesriverÎncă nu există evaluări
- ЧерепDocument34 paginiЧерепbekafop813Încă nu există evaluări
- Cochlear Anatomy: January 2008Document15 paginiCochlear Anatomy: January 2008Sekar ArumÎncă nu există evaluări
- Functional Anatomy of SkullDocument83 paginiFunctional Anatomy of SkullTaha Ahsan100% (1)
- Anatomy of The SkullDocument3 paginiAnatomy of The SkullYeraldin EspañaÎncă nu există evaluări
- 06 Lec 11 12 Anatomy First YearDocument13 pagini06 Lec 11 12 Anatomy First YearS:M:EÎncă nu există evaluări
- Anatomy of Petrous FaceDocument15 paginiAnatomy of Petrous FaceKelompok F8 CoasÎncă nu există evaluări
- Lec 6 Anatomy First StageDocument36 paginiLec 6 Anatomy First Stagespo9equtabaÎncă nu există evaluări
- Bones of The H&NDocument12 paginiBones of The H&Naminshafihassan902Încă nu există evaluări
- Enfermedad Vestibular FelinaDocument8 paginiEnfermedad Vestibular FelinaPamela Susana Rivera GómezÎncă nu există evaluări
- Surgical Anatomy of Mandible: Department of Oral and Maxilofacial SurgeryDocument92 paginiSurgical Anatomy of Mandible: Department of Oral and Maxilofacial SurgerySAMEENAÎncă nu există evaluări
- Rhoton - Cochlear NucleiDocument12 paginiRhoton - Cochlear NucleicristianescÎncă nu există evaluări
- Found 903128372 4535Document2 paginiFound 903128372 4535Bogdan Robert CÎncă nu există evaluări
- Brainstem: Medulla OblongataDocument130 paginiBrainstem: Medulla OblongataBhawana BhatodiyaÎncă nu există evaluări
- ანატომია ზეპირიDocument22 paginiანატომია ზეპირიmaÎncă nu există evaluări
- The Bones in The Neurocranium: David Terfera Shereen Jegtvig Clinical Anatomy For DummiesDocument25 paginiThe Bones in The Neurocranium: David Terfera Shereen Jegtvig Clinical Anatomy For DummiesFrozen Pandora MahayaÎncă nu există evaluări
- Anatomy Final All Questions (New)Document175 paginiAnatomy Final All Questions (New)Sevval OzcelikÎncă nu există evaluări
- Right Modified Neck Dissection (MND) : Case Study ONDocument71 paginiRight Modified Neck Dissection (MND) : Case Study ONAUSTIN SABUÎncă nu există evaluări
- Lecture Medskull - ModofiedDocument88 paginiLecture Medskull - ModofiedNikolai SamaniegoÎncă nu există evaluări
- Neuroanatomy Revision Notes PDFDocument7 paginiNeuroanatomy Revision Notes PDFTanya Tanu100% (1)
- The Infratemporal FossaDocument5 paginiThe Infratemporal FossaxxyumeÎncă nu există evaluări
- Cranial Nerves V, Ix, X, XiDocument89 paginiCranial Nerves V, Ix, X, XiUloko Christopher100% (1)
- Fetal Skull - Madam JustinaDocument11 paginiFetal Skull - Madam JustinaNana Yunus67% (3)
- The SkullDocument28 paginiThe SkullAyethÎncă nu există evaluări
- Basis CraniiDocument11 paginiBasis Craniicharlesy TÎncă nu există evaluări
- Anatomy AssignmentDocument10 paginiAnatomy AssignmentObansa JohnÎncă nu există evaluări
- Aa AnatomyDocument109 paginiAa AnatomyshailendraÎncă nu există evaluări
- 1516253699FSC P11 M4 E-TextDocument14 pagini1516253699FSC P11 M4 E-TextKanchanÎncă nu există evaluări
- Interior of The SkulDocument14 paginiInterior of The SkulArshad hussainÎncă nu există evaluări
- Introduction To Cranial CavityDocument35 paginiIntroduction To Cranial CavityexoÎncă nu există evaluări
- Found 903118732 4683Document2 paginiFound 903118732 4683Bogdan Robert CÎncă nu există evaluări
- Anatomy and Development of Oral Cavity and PharynxDocument14 paginiAnatomy and Development of Oral Cavity and PharynxGabriela MacoveanuÎncă nu există evaluări
- GRDA Intro NeckDocument12 paginiGRDA Intro NeckKingÎncă nu există evaluări
- Anatomy of The External EarDocument28 paginiAnatomy of The External EarhamaÎncă nu există evaluări
- Pharynx Anatomy - Overview, Gross Anatomy, Microscopic AnatomyDocument9 paginiPharynx Anatomy - Overview, Gross Anatomy, Microscopic AnatomyCarmen-BadeaÎncă nu există evaluări
- Chapter 10: Anatomy of The Pharynx and Esophagus P. Beasley Embryological DevelopmentDocument33 paginiChapter 10: Anatomy of The Pharynx and Esophagus P. Beasley Embryological DevelopmentGirish SubashÎncă nu există evaluări
- Middle Cranial FossaDocument5 paginiMiddle Cranial Fossaalexandra663Încă nu există evaluări
- Cranial CavityDocument94 paginiCranial CavityShashank Tripathi100% (1)
- 2 Head and Neck Anatomy SPTH 204Document79 pagini2 Head and Neck Anatomy SPTH 204Noor KhalilÎncă nu există evaluări
- Roberts, Stephen, BeightonDocument10 paginiRoberts, Stephen, BeightonJulio AbarzuaÎncă nu există evaluări
- 2Document13 pagini2Elena PruniciÎncă nu există evaluări
- Treatment Timing and Multidisciplinary Approach in Apert SyndromeDocument6 paginiTreatment Timing and Multidisciplinary Approach in Apert SyndromeJulio AbarzuaÎncă nu există evaluări
- Roberts, Stephen, BeightonDocument10 paginiRoberts, Stephen, BeightonJulio AbarzuaÎncă nu există evaluări
- Roberts, Stephen, BeightonDocument10 paginiRoberts, Stephen, BeightonJulio AbarzuaÎncă nu există evaluări
- Caring For Infants and Their Families: CraniosynostosisDocument13 paginiCaring For Infants and Their Families: CraniosynostosisJulio AbarzuaÎncă nu există evaluări
- Caring For Infants and Their Families: CraniosynostosisDocument13 paginiCaring For Infants and Their Families: CraniosynostosisJulio AbarzuaÎncă nu există evaluări
- Embriologia Humana y Biologia Del Desarrollo CarlsonDocument14 paginiEmbriologia Humana y Biologia Del Desarrollo CarlsonJulio AbarzuaÎncă nu există evaluări
- 11Document9 pagini11Din PcÎncă nu există evaluări
- t3 Som 2014 1Document8 paginit3 Som 2014 1Julio AbarzuaÎncă nu există evaluări
- t2 Cendekiawan 2010 PDFDocument9 paginit2 Cendekiawan 2010 PDFJulio AbarzuaÎncă nu există evaluări
- t3 Som 2014 2Document14 paginit3 Som 2014 2Julio AbarzuaÎncă nu există evaluări
- Va Medica: Atypical Swallowing: A ReviewDocument11 paginiVa Medica: Atypical Swallowing: A ReviewJulio AbarzuaÎncă nu există evaluări
- Topical Review: Sleep Bruxism, Headaches, and Sleep-Disordered Breathing in Children and AdolescentsDocument10 paginiTopical Review: Sleep Bruxism, Headaches, and Sleep-Disordered Breathing in Children and AdolescentsJulio AbarzuaÎncă nu există evaluări
- Frazier-Bowers Et Al, 2016Document11 paginiFrazier-Bowers Et Al, 2016Julio AbarzuaÎncă nu există evaluări
- Sajnani, 2015Document8 paginiSajnani, 2015Julio AbarzuaÎncă nu există evaluări
- Björk Facial Growth in Man, Studied With The Aid of Metallic ImplantsDocument6 paginiBjörk Facial Growth in Man, Studied With The Aid of Metallic ImplantsNataly ComettaÎncă nu există evaluări
- Sajnani, 2015Document8 paginiSajnani, 2015Julio AbarzuaÎncă nu există evaluări
- The Gerson TherapyDocument43 paginiThe Gerson Therapyboboacer100% (1)
- Sajnani, 2015Document7 paginiSajnani, 2015Julio AbarzuaÎncă nu există evaluări
- Andersson El Al%2ssssssc 2012Document8 paginiAndersson El Al%2ssssssc 2012Julio AbarzuaÎncă nu există evaluări
- Sajnani, 2015Document7 paginiSajnani, 2015Julio AbarzuaÎncă nu există evaluări
- 1820 Celestial EventDocument8 pagini1820 Celestial EventDoor Of ElÎncă nu există evaluări
- High Court Judgment On Ex Party DecreeDocument2 paginiHigh Court Judgment On Ex Party Decreeprashant pathakÎncă nu există evaluări
- Ethics - FinalsDocument18 paginiEthics - Finalsannie lalangÎncă nu există evaluări
- Investigative Project Group 8Document7 paginiInvestigative Project Group 8Riordan MoraldeÎncă nu există evaluări
- Villa VeronicaDocument12 paginiVilla Veronicacj fontzÎncă nu există evaluări
- Chap 9 Special Rules of Court On ADR Ver 1 PDFDocument8 paginiChap 9 Special Rules of Court On ADR Ver 1 PDFambahomoÎncă nu există evaluări
- 2.2 Push and Pull Sources of InnovationDocument16 pagini2.2 Push and Pull Sources of Innovationbclarke113Încă nu există evaluări
- Matlab/Simulink Models For Typical Soft Starting Means For A DC MotorDocument6 paginiMatlab/Simulink Models For Typical Soft Starting Means For A DC MotorkensesanÎncă nu există evaluări
- Worcester Vs Ocampo - DigestDocument1 paginăWorcester Vs Ocampo - DigestMaria Raisa Helga YsaacÎncă nu există evaluări
- HeavyReding ReportDocument96 paginiHeavyReding ReportshethÎncă nu există evaluări
- Wallen Et Al-2006-Australian Occupational Therapy JournalDocument1 paginăWallen Et Al-2006-Australian Occupational Therapy Journal胡知行Încă nu există evaluări
- Research in NursingDocument54 paginiResearch in Nursingrockycamaligan2356Încă nu există evaluări
- Apcr MCR 3Document13 paginiApcr MCR 3metteoroÎncă nu există evaluări
- All New Keys DictionaryDocument7 paginiAll New Keys DictionaryvishntÎncă nu există evaluări
- Mahabharata Reader Volume 1 - 20062023 - Free SampleDocument107 paginiMahabharata Reader Volume 1 - 20062023 - Free SampleDileep GautamÎncă nu există evaluări
- Songs of KabirDocument342 paginiSongs of KabirSant MatÎncă nu există evaluări
- Asphalt Hot Mix Training ManualDocument91 paginiAsphalt Hot Mix Training Manualyazqa50% (2)
- Tips On Being A Successful StudentDocument2 paginiTips On Being A Successful Studentshimbir100% (3)
- Qsen CurriculumDocument5 paginiQsen Curriculumapi-280981631Încă nu există evaluări
- Operate A Word Processing Application BasicDocument46 paginiOperate A Word Processing Application Basicapi-24787158267% (3)
- Part 4 Basic ConsolidationDocument3 paginiPart 4 Basic Consolidationtαtmαn dє grєαtÎncă nu există evaluări
- Elc650 Ws Guidelines (250219)Document3 paginiElc650 Ws Guidelines (250219)panda_yien100% (1)
- ABHI Network List As On 30-06-2023Document3.401 paginiABHI Network List As On 30-06-20233uifbcsktÎncă nu există evaluări
- Remo Vs DevanaderaDocument2 paginiRemo Vs DevanaderaZymon Andrew MaquintoÎncă nu există evaluări
- ADDICTED (The Novel) Book 1 - The Original English TranslationDocument1.788 paginiADDICTED (The Novel) Book 1 - The Original English TranslationMónica M. Giraldo100% (7)
- Mission Veng 29th, 2019Document4 paginiMission Veng 29th, 2019Lasky ChhakchhuakÎncă nu există evaluări
- Business Intelligence in RetailDocument21 paginiBusiness Intelligence in RetailGaurav Kumar100% (1)
- Sancticity AllDocument21 paginiSancticity AllJames DeHart0% (1)
- Human Right and Humanitarian. by Solicitor KaturaDocument12 paginiHuman Right and Humanitarian. by Solicitor KaturaFlavian PangahÎncă nu există evaluări
- Edu 536 - Task A2 - pld5Document3 paginiEdu 536 - Task A2 - pld5api-281740174Încă nu există evaluări