S-ar putea să vă placă și
- Como agem Inseticidas: NeurotransmissoresDocument36 paginiComo agem Inseticidas: NeurotransmissoresAndréFerraz100% (1)
- Ação Dos Inseticidas Nos InsetosDocument36 paginiAção Dos Inseticidas Nos InsetosEvandro KellerÎncă nu există evaluări
- Controle químico de plantas daninhasDocument54 paginiControle químico de plantas daninhasFernando De Medeiros Ledesma0% (1)
- Mecanismo de Ação Dos InseticidasDocument7 paginiMecanismo de Ação Dos InseticidasMarcos Vinícios Ferreira RochaÎncă nu există evaluări
- Controle Químico de Doenças AgroDocument37 paginiControle Químico de Doenças AgroJorge Luis GiongoÎncă nu există evaluări
- Ap - Uso de Bioestimulantes Na Indução de Resistência para o Manejo FitossanitárioDocument23 paginiAp - Uso de Bioestimulantes Na Indução de Resistência para o Manejo FitossanitárioAnna Carolina BarbozaÎncă nu există evaluări
- Apostila InseticidasDocument35 paginiApostila Inseticidastiago linsÎncă nu există evaluări
- Toxicologia E Classificação Dos Inseticidas: Entomologia AgrícolaDocument72 paginiToxicologia E Classificação Dos Inseticidas: Entomologia AgrícolaGustavo SantosÎncă nu există evaluări
- Apostila HerbicidasDocument17 paginiApostila HerbicidasHerison CaetanoÎncă nu există evaluări
- Reguladores de Crescimento FV AvancadaDocument37 paginiReguladores de Crescimento FV AvancadaLuiz Felipe SiqueiraÎncă nu există evaluări
- Apostila AlgasDocument27 paginiApostila Algasmaria grazielaÎncă nu există evaluări
- Texto 07 Tabela Das Principais Diferencas Entre Plantas C3 C4 e Cam 2005 PDFDocument2 paginiTexto 07 Tabela Das Principais Diferencas Entre Plantas C3 C4 e Cam 2005 PDFBambi PeckÎncă nu există evaluări
- Ecotoxicologia dos Defensivos AgrícolasDocument36 paginiEcotoxicologia dos Defensivos AgrícolasClaudia AnjosÎncă nu există evaluări
- Proteção de Plantas com FungicidasDocument200 paginiProteção de Plantas com FungicidasGuilherme SouzaÎncă nu există evaluări
- Estrobilurina Antonio PDFDocument66 paginiEstrobilurina Antonio PDFJosé Carlos da Costa Jr.Încă nu există evaluări
- BIANCA Controle BiológicoDocument64 paginiBIANCA Controle BiológicoJhonny EvertonÎncă nu există evaluări
- Mip MilhoDocument89 paginiMip MilhoReal AgricolaÎncă nu există evaluări
- Plantas daninhas: características e impactosDocument69 paginiPlantas daninhas: características e impactosRaniele100% (1)
- Resistência A HerbicidasDocument33 paginiResistência A HerbicidasNeto100% (3)
- CarboxamidasDocument30 paginiCarboxamidasgarcez2002100% (3)
- Fungicidas registrados para videiraDocument4 paginiFungicidas registrados para videiraGustavoPrzybylskiÎncă nu există evaluări
- Morfologia Vegetal: Tecidos e Estruturas das PlantasDocument68 paginiMorfologia Vegetal: Tecidos e Estruturas das Plantasmerco1717Încă nu există evaluări
- Planilha de Custos de Mecanização Agrícola PDFDocument2 paginiPlanilha de Custos de Mecanização Agrícola PDFk-us8263Încă nu există evaluări
- A relação entre produção, translocação e uso de fotoassimilados em células vegetaisDocument39 paginiA relação entre produção, translocação e uso de fotoassimilados em células vegetaisDelivelton Alexandre LongarayÎncă nu există evaluări
- Fitotoxicidade de Herbicidas na cultura milho: interações entre herbicidas, épocas e métodos de aplicação e manejo da adubação nitrogenadaDe la EverandFitotoxicidade de Herbicidas na cultura milho: interações entre herbicidas, épocas e métodos de aplicação e manejo da adubação nitrogenadaÎncă nu există evaluări
- Disciplina Controle BiologicoDocument3 paginiDisciplina Controle BiologicojoaopauloramosdemeloÎncă nu există evaluări
- Comportamento Dos Herbicidas No Solo LivroDocument42 paginiComportamento Dos Herbicidas No Solo Livroelizeub2078Încă nu există evaluări
- Manchas Foliares, Míldios, Oídios, FerrugemDocument40 paginiManchas Foliares, Míldios, Oídios, Ferrugemjrturra100% (1)
- Identifique os principais percevejos da sojaDocument44 paginiIdentifique os principais percevejos da sojaWashington Cardoso da CostaÎncă nu există evaluări
- Principais herbicidas pré-emergentes para milhoDocument1 paginăPrincipais herbicidas pré-emergentes para milhoHelber M. dos ReisÎncă nu există evaluări
- Efeito de carryover de trifluralin e S-metolachlor em culturas de soja, feijão e milhoDocument8 paginiEfeito de carryover de trifluralin e S-metolachlor em culturas de soja, feijão e milhoagroufpiÎncă nu există evaluări
- Manual Cascudinhos CompletoDocument21 paginiManual Cascudinhos CompletoGlenio BruckÎncă nu există evaluări
- FungicidasDocument89 paginiFungicidasEduardo PortoÎncă nu există evaluări
- Tecnologia de Aplicação Turma XDocument23 paginiTecnologia de Aplicação Turma XkriativaÎncă nu există evaluări
- MECANISMOS+DE+AÇAO+DE+HERBICIDAS+02 UnlockedDocument52 paginiMECANISMOS+DE+AÇAO+DE+HERBICIDAS+02 UnlockedRafael FerreiraÎncă nu există evaluări
- EPS - Experimentos em Parcelas SubdivididasDocument71 paginiEPS - Experimentos em Parcelas SubdivididasAllan Reis50% (2)
- Doenças da soja: Nematoide de cisto e mofo brancoDocument20 paginiDoenças da soja: Nematoide de cisto e mofo brancoJoão LivinoÎncă nu există evaluări
- Guia Tecnico MilhoDocument100 paginiGuia Tecnico MilhoJanderMouraÎncă nu există evaluări
- Morfologia Externa de Raízes e CaulesDocument54 paginiMorfologia Externa de Raízes e CaulesFranciele Salvador100% (2)
- Controle químico de doenças de plantasDocument87 paginiControle químico de doenças de plantasLeonardo Zimmer Nascimento100% (1)
- Mimetizadores de AuxinaDocument15 paginiMimetizadores de AuxinaGabriel Mota OliveiraÎncă nu există evaluări
- Modos de ação de fungicidas e controle de doenças em culturasDocument19 paginiModos de ação de fungicidas e controle de doenças em culturasDaniela PezziniÎncă nu există evaluări
- Guia de Identificação de Insetos-Praga Na Cultura Do Trigo L Conhecimento AgronômicoDocument32 paginiGuia de Identificação de Insetos-Praga Na Cultura Do Trigo L Conhecimento AgronômicoMatheus Gabriel Ferreira100% (1)
- Aminoácidos e bioestimulantes para redução de estresses nas plantasDocument50 paginiAminoácidos e bioestimulantes para redução de estresses nas plantasTiago TommÎncă nu există evaluări
- Desenvolvimento da soja R1-R4Document10 paginiDesenvolvimento da soja R1-R4k-us8263Încă nu există evaluări
- Comportamento e interação de herbicidas com o solo na cultura da cana-de-açúcarDocument90 paginiComportamento e interação de herbicidas com o solo na cultura da cana-de-açúcarangelolopes11Încă nu există evaluări
- Diagnose sintomas fitotoxicidade herbicidasDocument63 paginiDiagnose sintomas fitotoxicidade herbicidasJuliano LorenzettiÎncă nu există evaluări
- Cálculo Adicionar K para Saturar CTC - GISMONTIDocument2 paginiCálculo Adicionar K para Saturar CTC - GISMONTIPaulo Sérgio dos Santos JuniorÎncă nu există evaluări
- Seletividade de defensivos à inimigos naturaisDocument10 paginiSeletividade de defensivos à inimigos naturaisBruna AssisÎncă nu există evaluări
- 1 HerbicidasinibidoresdeEPsPsDocument35 pagini1 HerbicidasinibidoresdeEPsPsAndré MaylaÎncă nu există evaluări
- Mini-Curso Herbicidas Parte-2 PDFDocument50 paginiMini-Curso Herbicidas Parte-2 PDFGabriel Mota OliveiraÎncă nu există evaluări
- Regulação hormonal vegetalDocument26 paginiRegulação hormonal vegetalJamylle CatharineÎncă nu există evaluări
- Controle Biológico No BrasilDocument103 paginiControle Biológico No BrasilThomas GottÎncă nu există evaluări
- Aula3 MorfoVegetal 2019 1Document22 paginiAula3 MorfoVegetal 2019 1JulianyÎncă nu există evaluări
- Mecanismos de ação de herbicidasDocument36 paginiMecanismos de ação de herbicidasMoema Pascoini100% (1)
- Herbicidas em GramadosDocument33 paginiHerbicidas em GramadosrafaelÎncă nu există evaluări
- Potencialidades Da Moringa Oleifera Lam. No Semiárido Nordestino BrasileiroDe la EverandPotencialidades Da Moringa Oleifera Lam. No Semiárido Nordestino BrasileiroÎncă nu există evaluări
- Adubação Nitrogenada da Cultura do Trigo: com Base na Clorofilometria Via Aeronave Remotamente PilotadaDe la EverandAdubação Nitrogenada da Cultura do Trigo: com Base na Clorofilometria Via Aeronave Remotamente PilotadaÎncă nu există evaluări
- Livro de ReceitasDocument25 paginiLivro de ReceitasIsabela Ferreira Dos SantosÎncă nu există evaluări
- Gastronomia JulhoDocument33 paginiGastronomia Julhofcostos100% (2)
- 050 GABO SuperiorDocument1 pagină050 GABO SuperiorLuiz Eduardo AbreuÎncă nu există evaluări
- Comunicado SEAGRIDocument1 paginăComunicado SEAGRILuiz Eduardo AbreuÎncă nu există evaluări
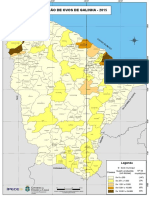
- Producao Ovos 2015Document1 paginăProducao Ovos 2015Luiz Eduardo AbreuÎncă nu există evaluări
- Chamada Oficial das Falanges MissionáriasDocument1 paginăChamada Oficial das Falanges MissionáriasLuiz Eduardo AbreuÎncă nu există evaluări
- Concurso Público para Assistente em Administração analisa dismorfia do SnapchatDocument10 paginiConcurso Público para Assistente em Administração analisa dismorfia do SnapchatIsadora TavaresÎncă nu există evaluări
- DC 054Document19 paginiDC 054Luiz Eduardo AbreuÎncă nu există evaluări
- PULGÕES, MOSCAS-BRANCAS E CIGARRINHAS Classificação de Inseticidas por MoADocument2 paginiPULGÕES, MOSCAS-BRANCAS E CIGARRINHAS Classificação de Inseticidas por MoAjordanaflores100% (1)
- Rel Doc NovoDocument3 paginiRel Doc NovoArlene PaixãoÎncă nu există evaluări
- Edital UfpeDocument11 paginiEdital UfpeKahMestreÎncă nu există evaluări
- AFRFB Simulado 1 - Pré-editalDocument55 paginiAFRFB Simulado 1 - Pré-editalLuiz Eduardo AbreuÎncă nu există evaluări
- 4 Dicas Incríveis de Como Fazer Um Abstract Conforme ABNTDocument8 pagini4 Dicas Incríveis de Como Fazer Um Abstract Conforme ABNTLuiz Eduardo AbreuÎncă nu există evaluări
- 9 Dicas Pré-Projeto MestradoDocument8 pagini9 Dicas Pré-Projeto MestradoLuiz Eduardo AbreuÎncă nu există evaluări
- 8 Dicas Artigo CientíficoDocument8 pagini8 Dicas Artigo CientíficoLuiz Eduardo AbreuÎncă nu există evaluări
- Matemática BásicaDocument68 paginiMatemática BásicaSantosÎncă nu există evaluări
- D9263 PNCFDocument2 paginiD9263 PNCFLuiz Eduardo AbreuÎncă nu există evaluări
- 7 Dicas de Como Fazer Uma Revisão Bibliográfica IncrívelDocument8 pagini7 Dicas de Como Fazer Uma Revisão Bibliográfica IncrívelLuiz Eduardo AbreuÎncă nu există evaluări
- Estudo Do Livro 10 Principios Basicos para Educar Seus FilhosDocument32 paginiEstudo Do Livro 10 Principios Basicos para Educar Seus FilhosVictorlud Lud100% (3)
- Ebook - BrunnoDocument40 paginiEbook - BrunnoLuiz Eduardo AbreuÎncă nu există evaluări
- 2004 - To Do Municipio e o Territorio RuralDocument64 pagini2004 - To Do Municipio e o Territorio RuralJoão Pedro Cilli DavidÎncă nu există evaluări
- Leis estaduais sobre APAC, denominações e alterações tributáriasDocument80 paginiLeis estaduais sobre APAC, denominações e alterações tributáriasvelhobianoÎncă nu există evaluări
- Cronograma Nivel Medio POS Edital COM LOGO PDFDocument9 paginiCronograma Nivel Medio POS Edital COM LOGO PDFLuiz Eduardo AbreuÎncă nu există evaluări
- Sesc PréDocument1 paginăSesc PréLuiz Eduardo AbreuÎncă nu există evaluări
- Estresse 1Document22 paginiEstresse 1Luiz Eduardo AbreuÎncă nu există evaluări
- Canteiro-Biossepticopdf PDFDocument28 paginiCanteiro-Biossepticopdf PDFLuiz Eduardo AbreuÎncă nu există evaluări
- Como ter educação financeiraDocument23 paginiComo ter educação financeiraLuiz Eduardo Abreu100% (1)
- Lei Ambiental Protege NaturezaDocument26 paginiLei Ambiental Protege NaturezaDêvis Furtado SouzaÎncă nu există evaluări
- CodornasDocument18 paginiCodornascarlostramonteÎncă nu există evaluări
- Plano de Aula 01 HandebolDocument2 paginiPlano de Aula 01 HandebolÍtalo Jorge Alves AlbuquerqueÎncă nu există evaluări
- CFTV Soluções SiemensDocument20 paginiCFTV Soluções SiemensTuga DolarÎncă nu există evaluări
- A Piramide MalditaDocument11 paginiA Piramide MalditaFelipe NunesÎncă nu există evaluări
- Mame Tutorial CompletoDocument43 paginiMame Tutorial CompletoCristien Brasso de CamposÎncă nu există evaluări
- Criando consultas SAP em 3 passosDocument4 paginiCriando consultas SAP em 3 passosjaimegrodriguesÎncă nu există evaluări
- Guia Do Linux - Avançado - SAMBA - Controle de Acesso Ao Servidor SAMBA - WikilivrosDocument10 paginiGuia Do Linux - Avançado - SAMBA - Controle de Acesso Ao Servidor SAMBA - Wikilivrossilviosergio2Încă nu există evaluări
- LOGIC Probabilidade Revisão EnemDocument4 paginiLOGIC Probabilidade Revisão EnemPedro RosaÎncă nu există evaluări
- Verbos na músicaDocument1 paginăVerbos na músicanarazevedo4337Încă nu există evaluări
- Áreas de Regiones CircularesDocument4 paginiÁreas de Regiones CircularesChristopher BlakeÎncă nu există evaluări
- Manual Equitacao Site FinalDocument154 paginiManual Equitacao Site FinalLuís LousadaÎncă nu există evaluări
- Aprenda divisão de forma rápida e divertidaDocument30 paginiAprenda divisão de forma rápida e divertidaLorena Vieira100% (1)
- Manual Pioneer MVH-078UB PDFDocument2 paginiManual Pioneer MVH-078UB PDFguirochacr0% (1)
- Alisson Viana - Campo Harmônico Maior (Amostra em PDFDocument13 paginiAlisson Viana - Campo Harmônico Maior (Amostra em PDFEd Master100% (1)
- Caixas Acústicas CSR 4000 - Manual de InstruçõesDocument22 paginiCaixas Acústicas CSR 4000 - Manual de InstruçõesSávio Gonçalves89% (9)
- (Tutorial PS2) Jogos de Fácil Tradução (MoH Van, Euro. Ass.Document8 pagini(Tutorial PS2) Jogos de Fácil Tradução (MoH Van, Euro. Ass.Jeremy JonesÎncă nu există evaluări
- Avaliação de Educação Fisica 4º AnoDocument4 paginiAvaliação de Educação Fisica 4º AnoAntonio CardosoÎncă nu există evaluări
- Fábula da Cigarra e da FormigaDocument1 paginăFábula da Cigarra e da FormigaPrMarcelo Fernanda AdonoÎncă nu există evaluări
- Regras de Jogo - Handebol Indoor 1º de Março de 2022Document103 paginiRegras de Jogo - Handebol Indoor 1º de Março de 2022Andressa XavierÎncă nu există evaluări
- A Dança Na Perspectiva de Um Jogo DigitalDocument24 paginiA Dança Na Perspectiva de Um Jogo DigitalCELINA DE BARROS BAIRÃOÎncă nu există evaluări
- Deum ErdDocument4 paginiDeum ErdEdimar N. MonteiroÎncă nu există evaluări
- A história de João do ValeDocument3 paginiA história de João do ValeDaniel FerreiraÎncă nu există evaluări
- Me AtraiuDocument2 paginiMe AtraiuMarcus CammpÎncă nu există evaluări
- Tempos compostos do modo subjetivoDocument1 paginăTempos compostos do modo subjetivoLuis ChinaiÎncă nu există evaluări
- LAURA 35/35, LAURA 35/35 F Laura 35 A, Laura 35 Af: Caldera Mural de Gas Gas-WandheizkesselDocument20 paginiLAURA 35/35, LAURA 35/35 F Laura 35 A, Laura 35 Af: Caldera Mural de Gas Gas-WandheizkesselIván CascudoÎncă nu există evaluări
- CNL ProvaEB Minha RuaDocument4 paginiCNL ProvaEB Minha RuaclaraveronicoÎncă nu există evaluări
- IOMAT MT Imprensa OficialDocument5 paginiIOMAT MT Imprensa Oficial̶V̶i̶v̶i̶a̶n̶e̶̶O̶l̶i̶v̶e̶i̶r̶a̶Încă nu există evaluări
- Formar discípulos: o alvo do líder de célulaDocument6 paginiFormar discípulos: o alvo do líder de célulaCongregacional Fruto Do EspiritoÎncă nu există evaluări
- Burro influencia comportamentoDocument2 paginiBurro influencia comportamentoGreice Correa83% (12)
- Decreto Nº 63.881 - CicloviasDocument7 paginiDecreto Nº 63.881 - Cicloviasaugusto mesquitaÎncă nu există evaluări
- Cronograma Informática Do Básico Ao AvançadoDocument2 paginiCronograma Informática Do Básico Ao Avançadofelipe costaÎncă nu există evaluări