S-ar putea să vă placă și
- Rafinesque Medical Flora Vol. 1 & 2 (p.337)Document660 paginiRafinesque Medical Flora Vol. 1 & 2 (p.337)Pasha Urr100% (1)
- 35 Plant Structure Growth and DevelopmentDocument62 pagini35 Plant Structure Growth and DevelopmentMedinaÎncă nu există evaluări
- Plant Tissues and OrgansDocument76 paginiPlant Tissues and Organsaldinette75% (4)
- Plant Tissue PDFDocument63 paginiPlant Tissue PDFJitendra Kumar100% (1)
- Make Wood VinegarDocument2 paginiMake Wood Vinegarjeej00Încă nu există evaluări
- List of Species in FBS 21 - Taxonomy of Forest PlantsDocument29 paginiList of Species in FBS 21 - Taxonomy of Forest PlantsCapale MC100% (1)
- CH 8 Transport in Plants Worksheet 1 AnswerDocument6 paginiCH 8 Transport in Plants Worksheet 1 Answermohammed mahdy100% (1)
- Lecture 4 Leaves and Stem AnatomyDocument55 paginiLecture 4 Leaves and Stem AnatomyMuhammad FadhilÎncă nu există evaluări
- Plant Structures and Physiology NotesDocument97 paginiPlant Structures and Physiology Notesapi-292966101Încă nu există evaluări
- Internal Morphology of RootsDocument5 paginiInternal Morphology of RootsMatthew Reyes RemegiaÎncă nu există evaluări
- Natsci002 Laboratory - Midterm, Worksheet 3 (Montesino)Document5 paginiNatsci002 Laboratory - Midterm, Worksheet 3 (Montesino)summersibalamontesinoÎncă nu există evaluări
- 02.1 - Morphology of Flowering PlantsDocument48 pagini02.1 - Morphology of Flowering Plantshgurmaita4321Încă nu există evaluări
- PL MorphDocument9 paginiPL Morphel vnÎncă nu există evaluări
- Botlec The Stems 4Document10 paginiBotlec The Stems 4Jeff MarianoÎncă nu există evaluări
- Plant Structure - Answers Not CompleteDocument3 paginiPlant Structure - Answers Not CompleteAadi MahajanÎncă nu există evaluări
- Plant Structure, Growth, and Development: RootsDocument9 paginiPlant Structure, Growth, and Development: RootsHerlin MabalaÎncă nu există evaluări
- BOLIGAO Exercise 17 Morphology of The StemDocument4 paginiBOLIGAO Exercise 17 Morphology of The StemJason Greyzon Boligao IndianaÎncă nu există evaluări
- Anatomy of Flowering PlantsDocument6 paginiAnatomy of Flowering PlantsAnonymous -Încă nu există evaluări
- Economic Botany: San Diego State UniversityDocument13 paginiEconomic Botany: San Diego State UniversityLester MolinaÎncă nu există evaluări
- Lab Rep 14finalDocument20 paginiLab Rep 14finalReign RosadaÎncă nu există evaluări
- Morphology of Flowering PlantsDocument48 paginiMorphology of Flowering PlantsAlakesh Coldplay KalitaÎncă nu există evaluări
- Chapter 9 Higher Plants - 2020Document109 paginiChapter 9 Higher Plants - 2020SARA LIM YEN ZI MoeÎncă nu există evaluări
- MeristemDocument25 paginiMeristemhazizi100% (3)
- Chapter MeristemsDocument5 paginiChapter Meristemsyoura4203Încă nu există evaluări
- Angiosperms Lab ActivityDocument11 paginiAngiosperms Lab ActivityHaris Khan100% (1)
- Plant AnatomyDocument63 paginiPlant AnatomyRabin Bajagain.Încă nu există evaluări
- Laboratory 1Document3 paginiLaboratory 1Rigel YunzalÎncă nu există evaluări
- RevisionDocument16 paginiRevisionMedinaÎncă nu există evaluări
- Stems Lesson 1: Stem FunctionsDocument11 paginiStems Lesson 1: Stem Functionskesyah rubidoÎncă nu există evaluări
- Plant Form and Function: School of Biotechnology, International UniversityDocument66 paginiPlant Form and Function: School of Biotechnology, International UniversityThu AnhÎncă nu există evaluări
- BIOL 100 Plant Form and FunctionDocument9 paginiBIOL 100 Plant Form and FunctionNiña Jean Tormis AldabaÎncă nu există evaluări
- Stomata and Others (Bakita Bujhe Nis)Document8 paginiStomata and Others (Bakita Bujhe Nis)PukuÎncă nu există evaluări
- Presentasi Bagian-Bagian Akar Biologi SMADocument41 paginiPresentasi Bagian-Bagian Akar Biologi SMAElisa Frederica SiburianÎncă nu există evaluări
- Exercise 14Document2 paginiExercise 14Mikhail LandichoÎncă nu există evaluări
- Week 1-A Review On Plant Structure and FunctionDocument85 paginiWeek 1-A Review On Plant Structure and FunctionSleeping Beauty100% (1)
- Terminology Used in Plant DescriptionDocument13 paginiTerminology Used in Plant DescriptionAli HamzaÎncă nu există evaluări
- Plant TissueDocument23 paginiPlant Tissuepewdiefannn3Încă nu există evaluări
- Plant Biology-Final SubmissionDocument7 paginiPlant Biology-Final SubmissionAr Deepti ManojÎncă nu există evaluări
- Chapter - 6 Anatomy of Flowering PlantsDocument6 paginiChapter - 6 Anatomy of Flowering PlantsPriyansh PatelÎncă nu există evaluări
- Structure of PlantsDocument17 paginiStructure of PlantsJillian Lao100% (5)
- BOT01Lec3 Part 1Document96 paginiBOT01Lec3 Part 1Jan Leo QuinioÎncă nu există evaluări
- Root Anatomy: RootsDocument6 paginiRoot Anatomy: RootsKim John NateÎncă nu există evaluări
- Bio 1 Module 3 4 PDFDocument101 paginiBio 1 Module 3 4 PDFJoyvy KidoÎncă nu există evaluări
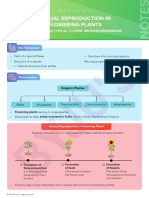
- CBSE Class 12 Sexual Reproduction in Flowering Plants Study Notes1Document80 paginiCBSE Class 12 Sexual Reproduction in Flowering Plants Study Notes1Fasil PattaraÎncă nu există evaluări
- Botany MidtermsDocument16 paginiBotany MidtermsAnisa AzisÎncă nu există evaluări
- Group 2-Excercise 2Document9 paginiGroup 2-Excercise 2James Carbonell Dela PeñaÎncă nu există evaluări
- Exercise No. 17Document9 paginiExercise No. 17Francis Jay MerencilloÎncă nu există evaluări
- Villanueva - Bot01 Lab Act #4Document14 paginiVillanueva - Bot01 Lab Act #4Kath ErineÎncă nu există evaluări
- 02 TissueDocument26 pagini02 Tissuecow dasÎncă nu există evaluări
- Makalah Bahasa Inggris: Giving Advice An Descriding PlantsDocument16 paginiMakalah Bahasa Inggris: Giving Advice An Descriding PlantsAyni bedduÎncă nu există evaluări
- Root Anatomy E-UnitDocument5 paginiRoot Anatomy E-UnitElaiza Aleman SisonÎncă nu există evaluări
- Lab Activity 5: Plant Morphology: StemDocument5 paginiLab Activity 5: Plant Morphology: StemPaulÎncă nu există evaluări
- Activity 11 - The Leaf (Instruction Sheet)Document9 paginiActivity 11 - The Leaf (Instruction Sheet)Chris P. BeaconÎncă nu există evaluări
- Group 2-Excercise 2Document10 paginiGroup 2-Excercise 2James Carbonell Dela PeñaÎncă nu există evaluări
- ROOTS ReviewerDocument5 paginiROOTS ReviewerMANALANG JULIA ALEXISÎncă nu există evaluări
- Exercise No. 7Document21 paginiExercise No. 7John MelbyÎncă nu există evaluări
- Chapter 8 Revisions III - QuizletDocument24 paginiChapter 8 Revisions III - QuizletChristianAvelinoÎncă nu există evaluări
- 11 Biology Notes Ch06 Anatomy of Flowering PlantsDocument8 pagini11 Biology Notes Ch06 Anatomy of Flowering PlantsPriyanshu BhadanaÎncă nu există evaluări
- Plant TissuesDocument6 paginiPlant TissuesJEVRIX RAMOSÎncă nu există evaluări
- Male Reproductive Organ Plants (Botany)Document23 paginiMale Reproductive Organ Plants (Botany)kunalÎncă nu există evaluări
- Xi-A of Atomic Energy Central School HasDocument27 paginiXi-A of Atomic Energy Central School Hasashok parmarÎncă nu există evaluări
- Wetland Plants of the Adirondacks: Ferns, Woody Plants, and GraminoidsDe la EverandWetland Plants of the Adirondacks: Ferns, Woody Plants, and GraminoidsÎncă nu există evaluări
- Wetland Plants of the Adirondacks: Herbaceous Plants and Aquatic PlantsDe la EverandWetland Plants of the Adirondacks: Herbaceous Plants and Aquatic PlantsÎncă nu există evaluări
- Mole Guided Notes PDFDocument2 paginiMole Guided Notes PDFapi-232487761Încă nu există evaluări
- Cookie Chemistry Lab2Document2 paginiCookie Chemistry Lab2api-232487761Încă nu există evaluări
- Cookie Chemistry Notes Fill in 2Document1 paginăCookie Chemistry Notes Fill in 2api-232487761Încă nu există evaluări
- Mitosis 2Document20 paginiMitosis 2api-232487761Încă nu există evaluări
- H9-Seed Dormancy 2013Document5 paginiH9-Seed Dormancy 2013api-232487761Încă nu există evaluări
- Seed Plants Lab: Learning ObjectivesDocument11 paginiSeed Plants Lab: Learning Objectivesapi-232487761Încă nu există evaluări
- Leaf Anatomy LabDocument3 paginiLeaf Anatomy Labapi-232487761Încă nu există evaluări
- Reebop-LabDocument2 paginiReebop-Labapi-232487761Încă nu există evaluări
- Rice Growth and DevelopmentDocument56 paginiRice Growth and DevelopmentSabin BhattaraiÎncă nu există evaluări
- After Completing This Module, You Are Expected To:: Learning ObjectivesDocument11 paginiAfter Completing This Module, You Are Expected To:: Learning ObjectivesRencel Joy Datoy100% (2)
- Hershey 1996Document9 paginiHershey 1996api-329324684Încă nu există evaluări
- Curry LeavesDocument3 paginiCurry Leaveswindow_7Încă nu există evaluări
- JCB Wheeled Loader 407bzx 4088 ZX 409b Z 410b ZX 411b ZX Sevice ManualDocument22 paginiJCB Wheeled Loader 407bzx 4088 ZX 409b Z 410b ZX 411b ZX Sevice Manualadamjoseph180384gbj100% (128)
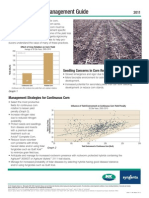
- NK SyngentaDocument2 paginiNK SyngentahansstevenÎncă nu există evaluări
- Fertilizer, Manure and Compost Effects On Weed Growth and Competition With Winter Wheat in Western CanadaDocument10 paginiFertilizer, Manure and Compost Effects On Weed Growth and Competition With Winter Wheat in Western CanadaLuis de los SantosÎncă nu există evaluări
- Ls 3 - Plant Kingdom: Algae, Bryophytes, Pteridophytes, Gymnosperms and AngiospermsDocument44 paginiLs 3 - Plant Kingdom: Algae, Bryophytes, Pteridophytes, Gymnosperms and AngiospermsShady WeebÎncă nu există evaluări
- Salad Mix Accomplishment ReportDocument2 paginiSalad Mix Accomplishment ReportRickxie GonzagaÎncă nu există evaluări
- Deciduous Forest 2P4Document15 paginiDeciduous Forest 2P4quekyk5029Încă nu există evaluări
- Plants and Society 8Th Edition Full ChapterDocument41 paginiPlants and Society 8Th Edition Full Chapterapril.benefiel764100% (27)
- Tle 6 - Periodic-ExamDocument8 paginiTle 6 - Periodic-ExamC VDÎncă nu există evaluări
- Module 1 Introduction To BotanyDocument11 paginiModule 1 Introduction To BotanyCEEJAY P. PATAWARANÎncă nu există evaluări
- Discusses The Importance of Planting and Propagating Trees and Fruit-Bearing Trees and Marketing SeedlingsDocument4 paginiDiscusses The Importance of Planting and Propagating Trees and Fruit-Bearing Trees and Marketing Seedlingscatherine renanteÎncă nu există evaluări
- DLL Tle-6 Q1 W4-JanDocument7 paginiDLL Tle-6 Q1 W4-JanJanille Tomajin-Capinpin100% (1)
- Pollination in Plants - Types, Advantages and Disadvantages PDFDocument28 paginiPollination in Plants - Types, Advantages and Disadvantages PDFRaj KumarÎncă nu există evaluări
- Calendar of Agricultural ActivitiesDocument2 paginiCalendar of Agricultural ActivitiesGenevieve MorilloÎncă nu există evaluări
- M20 Davi4493 08 Se C20Document66 paginiM20 Davi4493 08 Se C20Dbalt100% (1)
- Organic and Inorganic Fertilizers PDFDocument2 paginiOrganic and Inorganic Fertilizers PDFJeff Raian Molina BatongbakalÎncă nu există evaluări
- Covo Rugare Gold PLUSDocument1 paginăCovo Rugare Gold PLUSFarai50% (2)
- Plant Transport SystemsDocument20 paginiPlant Transport Systemscale suarezÎncă nu există evaluări
- Solution Assignment2Document1 paginăSolution Assignment2Daman RajÎncă nu există evaluări
- Morphoanatomical Profile of Five Species of Piper L. From Bangladesh and Its Taxonomic SignificanceDocument12 paginiMorphoanatomical Profile of Five Species of Piper L. From Bangladesh and Its Taxonomic SignificanceKhairunnisa Salsabila LutfiÎncă nu există evaluări
- Morphology of Flowering Plants BiohackDocument14 paginiMorphology of Flowering Plants BiohackRahul KUMAR Nayak100% (1)
- 5.5 Green Belt Development PlanDocument9 pagini5.5 Green Belt Development PlanratannsÎncă nu există evaluări
- Botany (UG & PG)Document1 paginăBotany (UG & PG)Harsha BabyÎncă nu există evaluări