S-ar putea să vă placă și
- Reporte #14Document1 paginăReporte #14Paola CasasÎncă nu există evaluări
- Trafico Intracelular-SulyDocument3 paginiTrafico Intracelular-SulySuly VigoÎncă nu există evaluări
- Ensayo de BioquimicaDocument3 paginiEnsayo de BioquimicaandreaÎncă nu există evaluări
- Perez Viñan Araceli de Los Angeles C2u1Document12 paginiPerez Viñan Araceli de Los Angeles C2u1Araceli PérezÎncă nu există evaluări
- 4.°sec - Biologíanm - Ibim2020 - Ficha de Reforzamiento 1Document5 pagini4.°sec - Biologíanm - Ibim2020 - Ficha de Reforzamiento 1Kiara BalcazarÎncă nu există evaluări
- Resumen Celula y Organelos KineDocument4 paginiResumen Celula y Organelos KineSam Dobson100% (1)
- La Celula y Su Fisiologia Trabajo ImprimirDocument9 paginiLa Celula y Su Fisiologia Trabajo Imprimircamarita1978Încă nu există evaluări
- Taller No. 1 Tejido EpitelialDocument11 paginiTaller No. 1 Tejido Epitelialangelly pinillaÎncă nu există evaluări
- Morfofunción Del Sistema DigestivoDocument110 paginiMorfofunción Del Sistema DigestivoSelena San AndrésÎncă nu există evaluări
- Sinapsis I 2012 PDFDocument27 paginiSinapsis I 2012 PDFleticiaÎncă nu există evaluări
- Categorias Taxonomicas de Los Seres VivosDocument20 paginiCategorias Taxonomicas de Los Seres VivosChristian Javier Mendoza LozadaÎncă nu există evaluări
- Chlamydia TrachomatisDocument35 paginiChlamydia TrachomatisLizet Judith Osorio GonzálezÎncă nu există evaluări
- Crucigrama BiomoleculasDocument1 paginăCrucigrama BiomoleculasAnónimo 88100% (1)
- Solucionario Guía Ciclo CelularDocument12 paginiSolucionario Guía Ciclo CelularAriel100% (1)
- s11 - PPT - Bartonella - Yersinia - Hongos.Document30 paginis11 - PPT - Bartonella - Yersinia - Hongos.dayanaÎncă nu există evaluări
- Álbum de HistologíaDocument59 paginiÁlbum de HistologíaClaudia Miranda CarreñoÎncă nu există evaluări
- Cuadro Sistema Endocrino Hormonas y Glandulas (Autoguardado)Document6 paginiCuadro Sistema Endocrino Hormonas y Glandulas (Autoguardado)Octavio Irazabal100% (1)
- Los Asquelmintos y Su DivisiónDocument2 paginiLos Asquelmintos y Su DivisiónDiana SaenzÎncă nu există evaluări
- Micro Cap 25Document11 paginiMicro Cap 25Hellen MendezÎncă nu există evaluări
- Señalización Celular y Transducción de SeñalesDocument15 paginiSeñalización Celular y Transducción de SeñalesJuan Felipe Salas Ospina100% (1)
- Practica 5 (Mitosis) - Biologia - P 108 - Macas CarlosDocument3 paginiPractica 5 (Mitosis) - Biologia - P 108 - Macas CarlosCarlosMacasÎncă nu există evaluări
- Actividad 2 Cuadro Comparativo Tipos de MusculosDocument2 paginiActividad 2 Cuadro Comparativo Tipos de MusculosVictor LorcaÎncă nu există evaluări
- Casos Clínicos de Bioquímica Molecular (Colera)Document6 paginiCasos Clínicos de Bioquímica Molecular (Colera)Criss2720Încă nu există evaluări
- Cuestionario de Sistema Endocrino, HistologíaDocument10 paginiCuestionario de Sistema Endocrino, HistologíaDayana IntriagoÎncă nu există evaluări
- Ensayo BiologiaDocument4 paginiEnsayo BiologiaLida Luz BeltranÎncă nu există evaluări
- Meiosis PDFDocument22 paginiMeiosis PDFDeyfer Steven Cortes Castrillon100% (1)
- Glosario Membrana CelularDocument3 paginiGlosario Membrana CelularNicole OrjuelaÎncă nu există evaluări
- 08 Guia Meiosis y Gametogenesis - 2016 - PRODocument16 pagini08 Guia Meiosis y Gametogenesis - 2016 - PROlordviricÎncă nu există evaluări
- Hongos.1Document56 paginiHongos.1Eder Javier Fernández Huamani0% (1)
- Sistema Inmune Glosario PDFDocument7 paginiSistema Inmune Glosario PDFCAMILA VARGASÎncă nu există evaluări
- Carioteca DiaposibibaDocument6 paginiCarioteca DiaposibibaCristhian DraxlerÎncă nu există evaluări
- Actividad 3. Órganos Primarios y Secundarios Del Sistema InmuneDocument10 paginiActividad 3. Órganos Primarios y Secundarios Del Sistema InmuneSarai GomezÎncă nu există evaluări
- Citoesqueleto ForoDocument39 paginiCitoesqueleto ForoClaudia Montes GuerraÎncă nu există evaluări
- Capítulo 28 Los Aparatos ReproductoresDocument5 paginiCapítulo 28 Los Aparatos ReproductoresAbraham RamirezÎncă nu există evaluări
- Membrana Plasmática PDFDocument37 paginiMembrana Plasmática PDFErika ToponÎncă nu există evaluări
- Semana 11 - Biología Celular - Señalización CelularDocument10 paginiSemana 11 - Biología Celular - Señalización CelularSofía HerradaÎncă nu există evaluări
- Unidad y Diversidad de Las CélulasDocument6 paginiUnidad y Diversidad de Las CélulasRita Maribel Bonilla Mora0% (1)
- Regulacion CalcemiaDocument69 paginiRegulacion CalcemiaIrlanda Maldonado100% (1)
- Preguntas de Tejidos BasicosDocument2 paginiPreguntas de Tejidos BasicosDayana AguirreÎncă nu există evaluări
- Solucion de Bioquimica GenaralDocument4 paginiSolucion de Bioquimica GenaralElsa Santos tocasÎncă nu există evaluări
- PROTEOSOMASDocument11 paginiPROTEOSOMASMariana Palacio100% (1)
- Mapa Conceptual Integradora 3Document2 paginiMapa Conceptual Integradora 3Francisco Javier Quiroz0% (1)
- Cuestionario "La Célula"Document5 paginiCuestionario "La Célula"MsQBPÎncă nu există evaluări
- Glosario de TerminosDocument5 paginiGlosario de TerminosMaria plataÎncă nu există evaluări
- Flagelos OficialDocument21 paginiFlagelos OficialMarisol BravoÎncă nu există evaluări
- AntibioticosDocument1 paginăAntibioticosKELLY DAYAN GARCIA JIMENEZÎncă nu există evaluări
- Preguntas Sobre Agua y Sales MineralesDocument3 paginiPreguntas Sobre Agua y Sales MineralesOliver Abreu CepedaÎncă nu există evaluări
- Carbohidatos 1Document1 paginăCarbohidatos 1Majo ReyesÎncă nu există evaluări
- Embriogenesis de La Cuarta Semana A La Octava SemanaDocument55 paginiEmbriogenesis de La Cuarta Semana A La Octava SemanaZOfii SnhzÎncă nu există evaluări
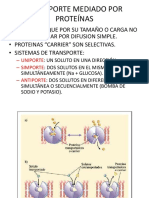
- Transporte Mediado Por ProteínasDocument8 paginiTransporte Mediado Por ProteínasMaite Celi LoayzaÎncă nu există evaluări
- Resumen Capitulo 2 AbbasDocument4 paginiResumen Capitulo 2 AbbasAngie TatianaÎncă nu există evaluări
- Tabla de OrganelosDocument2 paginiTabla de OrganelosIvette Herrera100% (1)
- Cuestionario Cinética EnzimáticaDocument2 paginiCuestionario Cinética EnzimáticaIsabel MartinezÎncă nu există evaluări
- Estructura de La MitocondriaDocument2 paginiEstructura de La MitocondriasaimyÎncă nu există evaluări
- Caso Problema Guion DefinitivoDocument9 paginiCaso Problema Guion Definitivocarolina arcosÎncă nu există evaluări
- Transporte Vesicular Aparato de GolgiDocument54 paginiTransporte Vesicular Aparato de Golgiimarosen50% (2)
- Los Cromosomas (Cuadro Comparativo)Document2 paginiLos Cromosomas (Cuadro Comparativo)crimes100% (1)
- Nombre: Astrid Zapata: Taller N°2Document8 paginiNombre: Astrid Zapata: Taller N°2Ken Sants100% (1)
- Tema Ii. Potencial de Membrana o Potencial de AcciónDocument8 paginiTema Ii. Potencial de Membrana o Potencial de AcciónCristina Jonas LopezÎncă nu există evaluări
- Tema 3 DesarrolloDocument16 paginiTema 3 Desarrolloantonio garcia martinezÎncă nu există evaluări
- Manual de PediatríaDocument104 paginiManual de Pediatríadanitza pilco100% (6)
- Manual de OtorrinolaringologíaDocument68 paginiManual de Otorrinolaringologíadanitza pilco100% (2)
- Manual de PsiquiatríaDocument84 paginiManual de Psiquiatríadanitza pilco87% (15)
- Manual de Neumología y Cirugía Torácica PDFDocument108 paginiManual de Neumología y Cirugía Torácica PDFdanitza pilcoÎncă nu există evaluări
- Manual de NefrologíaDocument84 paginiManual de Nefrologíadanitza pilcoÎncă nu există evaluări
- Manual de InmunologíaDocument44 paginiManual de Inmunologíadanitza pilco100% (1)
- Manual de Neurología y Neurocirugía PDFDocument120 paginiManual de Neurología y Neurocirugía PDFSolcito De verano100% (5)
- Manual de Digestivo y Cirugía General PDFDocument188 paginiManual de Digestivo y Cirugía General PDFRolando Gtmero100% (1)
- Manual de EndocrinologíaDocument108 paginiManual de Endocrinologíadanitza pilco100% (5)
- Libro Gordo 2009 - 2019Document824 paginiLibro Gordo 2009 - 2019Sofia Yate Ochoa100% (7)
- Ginecología PDFDocument120 paginiGinecología PDFSebas Rodriguez100% (2)
- Manual de Infecciosas y Microbiología PDFDocument156 paginiManual de Infecciosas y Microbiología PDFdanitza pilco100% (1)
- Manual de Traumatología y Cirugía OrtopédicaDocument84 paginiManual de Traumatología y Cirugía Ortopédicadanitza pilco100% (8)
- Reumatologia 11ed-2019Document116 paginiReumatologia 11ed-2019danitza pilco100% (5)
- Manual de Dermatología 12 EdiciónDocument72 paginiManual de Dermatología 12 EdiciónALFREDO JUNIOR LEON ALEJO100% (6)
- Estadística PDFDocument56 paginiEstadística PDFSebas Rodriguez100% (1)
- AMIR España 12 - CardiologíaDocument148 paginiAMIR España 12 - Cardiologíaedison aruquipa100% (11)
- Estadistica y Epidemiologia 11ed-2019 PDFDocument66 paginiEstadistica y Epidemiologia 11ed-2019 PDFdanitza pilco100% (1)
- Manual de Hematología PDFDocument92 paginiManual de Hematología PDFdanitza pilco67% (3)
- Otorrinolaringologia 11ed-2019Document98 paginiOtorrinolaringologia 11ed-2019danitza pilco67% (3)
- Psiquiatria 11ed-2019 PDFDocument106 paginiPsiquiatria 11ed-2019 PDFdanitza pilco100% (2)
- Manual de OtorrinolaringologíaDocument68 paginiManual de Otorrinolaringologíadanitza pilco100% (2)
- Manual de UrologíaDocument76 paginiManual de Urologíadanitza pilco67% (3)
- Manual de EndocrinologíaDocument108 paginiManual de Endocrinologíadanitza pilco100% (5)
- Manual de PsiquiatríaDocument84 paginiManual de Psiquiatríadanitza pilco87% (15)
- Manual de NefrologíaDocument84 paginiManual de Nefrologíadanitza pilcoÎncă nu există evaluări
- EdafologíaDocument35 paginiEdafologíaapi-3702178100% (9)
- Funciones de Los Polisacaridos CelulosaDocument2 paginiFunciones de Los Polisacaridos CelulosaConstanza Coronado RiveraÎncă nu există evaluări
- Tema 1 Glandulas Salivales PDFDocument66 paginiTema 1 Glandulas Salivales PDFMIGUEL ANGEL CALLE GRANADAÎncă nu există evaluări
- Antihipertensivos2020 1Document22 paginiAntihipertensivos2020 1Arleny TaverasÎncă nu există evaluări
- Unidad I Fisicoquimica II 2020Document14 paginiUnidad I Fisicoquimica II 2020Antonela YanethÎncă nu există evaluări
- Solidos No CristalinosDocument11 paginiSolidos No CristalinosyuliÎncă nu există evaluări
- Rendimientos PDFDocument175 paginiRendimientos PDFA-Mishel RivasplataÎncă nu există evaluări
- Titulación Con IndicadorDocument22 paginiTitulación Con IndicadorDíaz Gutierrez AranzaÎncă nu există evaluări
- Hds Applaud-25-Wp Insecticida VerdeDocument7 paginiHds Applaud-25-Wp Insecticida VerdeFrancisca Madrid MadridÎncă nu există evaluări
- Estabilizacion Suelo - Cemento, Cal - Cal-Cemento - Emulsion 1Document47 paginiEstabilizacion Suelo - Cemento, Cal - Cal-Cemento - Emulsion 1Gabriela100% (1)
- Creolina-Triple FTDocument3 paginiCreolina-Triple FTdiego monte100% (1)
- Protocolo de CALIDADDocument39 paginiProtocolo de CALIDADMARIA JOSE ALEXANDRA AYALA OSORIOÎncă nu există evaluări
- COLORANTESDocument9 paginiCOLORANTESJaydy Chiroque VelizÎncă nu există evaluări
- Catalogo Jofel 2014-15Document36 paginiCatalogo Jofel 2014-15arqd7Încă nu există evaluări
- Trabajo Final AdsorcionDocument23 paginiTrabajo Final AdsorcionAlexa VelezÎncă nu există evaluări
- Tecnología Enzimática PDFDocument35 paginiTecnología Enzimática PDFcaruingÎncă nu există evaluări
- Sintesis de Compuestos InorganicosDocument16 paginiSintesis de Compuestos InorganicosChristian Obispo100% (1)
- Formulas Patron y PropuestaDocument4 paginiFormulas Patron y PropuestaYASMINE RINCON RELÎncă nu există evaluări
- Ejercicios de Electroquimica 43918Document2 paginiEjercicios de Electroquimica 43918LuisÎncă nu există evaluări
- Irrigacion en EndodonciaDocument10 paginiIrrigacion en EndodonciaAf BqÎncă nu există evaluări
- Final Cosmetologia ResumenDocument194 paginiFinal Cosmetologia ResumenguilleromopulidoÎncă nu există evaluări
- Diseño y Estructura de MotoresDocument31 paginiDiseño y Estructura de MotoresPercyRivera100% (1)
- AspartameDocument6 paginiAspartameDavid Huamán GastelúÎncă nu există evaluări
- La Plataforma Estable de Grebennikov v1.0Document11 paginiLa Plataforma Estable de Grebennikov v1.0TODO SOBRE OVNISÎncă nu există evaluări
- Mendiola Juan Soluciones ValoradasDocument7 paginiMendiola Juan Soluciones Valoradasany perez fonsecaÎncă nu există evaluări
- Promotor de Adherencia para PlásticosDocument2 paginiPromotor de Adherencia para PlásticosKing WAÎncă nu există evaluări
- Métodos Básicos para El Estudio de La Sensibilidad A Los AntimicrobianosDocument58 paginiMétodos Básicos para El Estudio de La Sensibilidad A Los AntimicrobianosDaniel Pacheco RojasÎncă nu există evaluări
- CARBOHIDRATOS Mollinedo PatziDocument4 paginiCARBOHIDRATOS Mollinedo PatziGenaro Chanco MendozaÎncă nu există evaluări