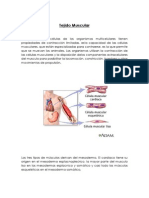
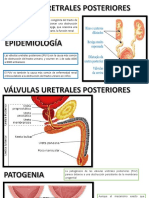
S-ar putea să vă placă și
- Musculo LisoDocument5 paginiMusculo LisoChicharitho HernándezÎncă nu există evaluări
- Músculo Liso ExpoDocument6 paginiMúsculo Liso ExpoTania CarmonaÎncă nu există evaluări
- Informe Histologia Tejido MuscularClaraDocument8 paginiInforme Histologia Tejido MuscularClaraLilibeth Fajardo MedinaÎncă nu există evaluări
- Evaluacion de La Contraccion MuscularDocument18 paginiEvaluacion de La Contraccion MuscularAriana OroscoÎncă nu există evaluări
- Guia Tejido MuscularDocument6 paginiGuia Tejido MuscularDiana Esmeralda De los SantosÎncă nu există evaluări
- Muscular y CardiovascularDocument20 paginiMuscular y CardiovascularOctavio GarroneÎncă nu există evaluări
- Taller 5 de FisioDocument6 paginiTaller 5 de FisioxiomararsÎncă nu există evaluări
- Esenciales MuscularDocument2 paginiEsenciales MuscularSofi BremeÎncă nu există evaluări
- Introduccion Musculo LisoDocument2 paginiIntroduccion Musculo LisoMiguel Cuellar50% (2)
- Capitulo 5 GanongDocument9 paginiCapitulo 5 GanongMeli MolinaÎncă nu există evaluări
- Tejido MuscularDocument6 paginiTejido MuscularYae KRÎncă nu există evaluări
- CitoesqueletoDocument8 paginiCitoesqueletoAlejandra GuaillasÎncă nu există evaluări
- Musculo LisoDocument9 paginiMusculo LisoMartinVelazcoÎncă nu există evaluări
- Capitulo 8Document3 paginiCapitulo 8mariaisabel_1807Încă nu există evaluări
- Musculo LisoDocument38 paginiMusculo LisoAnalú ViteÎncă nu există evaluări
- Histología General PDFDocument7 paginiHistología General PDFBeiruth De LeónÎncă nu există evaluări
- Histología General PDFDocument7 paginiHistología General PDFBeiruth De LeónÎncă nu există evaluări
- Tejido MuscularDocument13 paginiTejido MuscularCamila Agostina VillafañeÎncă nu există evaluări
- Brenda Tejido Muscular Por Dentro Del Titan ColosalDocument6 paginiBrenda Tejido Muscular Por Dentro Del Titan ColosalJesus Abel Quiñonez ValenzuelaÎncă nu există evaluări
- Histología Del Tejido MuscularDocument4 paginiHistología Del Tejido MuscularArianny CamacaroÎncă nu există evaluări
- Musculo LisoDocument10 paginiMusculo Lisohedwig21Încă nu există evaluări
- 9 Conferencia Tejido MuscularDocument101 pagini9 Conferencia Tejido MuscularHenry CondoriÎncă nu există evaluări
- Histologia Tejido Muscular ResumenDocument5 paginiHistologia Tejido Muscular ResumenValentina Ortiz VarasÎncă nu există evaluări
- HistoDocument10 paginiHistoMacarena GiraudoÎncă nu există evaluări
- Contracción Del Musculo Liso - FisiologíaDocument5 paginiContracción Del Musculo Liso - FisiologíaDara DosÎncă nu există evaluări
- Informe Tejido Musuclar FinalDocument10 paginiInforme Tejido Musuclar FinalsripiiÎncă nu există evaluări
- Células ContráctilesDocument10 paginiCélulas ContráctilesmacaÎncă nu există evaluări
- Expo MiosinaDocument10 paginiExpo MiosinaAnndres Daviid BermudezÎncă nu există evaluări
- Musculo CardiacoDocument2 paginiMusculo CardiacoAlani FloresÎncă nu există evaluări
- Docsity Resumen de Histologia de Ross 1Document27 paginiDocsity Resumen de Histologia de Ross 1Van NanaÎncă nu există evaluări
- Tejido MuscularDocument3 paginiTejido MuscularJubelis BalcazarÎncă nu există evaluări
- TEMA 4 Tejido MuscularDocument6 paginiTEMA 4 Tejido MuscularMireya Garcia NietoÎncă nu există evaluări
- Contracción Del Músculo LisoDocument4 paginiContracción Del Músculo LisoJulia dhrÎncă nu există evaluări
- FisiologiaDocument50 paginiFisiologiaElizabeth SaavedraÎncă nu există evaluări
- 59 Circ Musculo Liso VascularDocument9 pagini59 Circ Musculo Liso VascularCarlos VargasÎncă nu există evaluări
- Tejido Muscular Histologia Libro Atlas Garner Hia...Document36 paginiTejido Muscular Histologia Libro Atlas Garner Hia...Karolina I Maldonado UcÎncă nu există evaluări
- Tejido MuscularDocument4 paginiTejido MuscularAna MalaisiÎncă nu există evaluări
- Filamentos de ActinaDocument6 paginiFilamentos de ActinaAgustina UriarteÎncă nu există evaluări
- Contracción MuscularDocument13 paginiContracción MuscularMariana EspitiaÎncă nu există evaluări
- Contración Músculo LisoDocument5 paginiContración Músculo LisoDiego MartinezÎncă nu există evaluări
- 7 Observacion de Celulas Musculares PDFDocument37 pagini7 Observacion de Celulas Musculares PDFmartin mahomaÎncă nu există evaluări
- Tejido MuscularDocument4 paginiTejido MuscularSandra TorelliÎncă nu există evaluări
- Clasificación Del Tejido MuscularDocument13 paginiClasificación Del Tejido MuscularJUAN CARLOS ROBAYO BOTIVA72% (18)
- Citoesqueleto (Fisiologia)Document11 paginiCitoesqueleto (Fisiologia)Julio César Reátegui PachecoÎncă nu există evaluări
- Musculo LisoDocument42 paginiMusculo LisoAriana AmparoÎncă nu există evaluări
- Tema 5 AcabadoDocument10 paginiTema 5 AcabadoMagdalena Oñate SerranoÎncă nu există evaluări
- Universidad Privada San Juan BautistaDocument12 paginiUniversidad Privada San Juan BautistaJhan Carlos Escobar OreÎncă nu există evaluări
- 7 Tejido - Muscular Ok 2020 - 1Document11 pagini7 Tejido - Muscular Ok 2020 - 1Romi SÎncă nu există evaluări
- Tejido Muscular - Exposicion 2Document30 paginiTejido Muscular - Exposicion 2Jordy Steven Betancourt AvalosÎncă nu există evaluări
- TEMA 4 - Tejido MuscularDocument7 paginiTEMA 4 - Tejido MuscularGLORIA RMÎncă nu există evaluări
- Citoplasma, Citoesqueleto, RibosomasDocument26 paginiCitoplasma, Citoesqueleto, Ribosomas2cr_28Încă nu există evaluări
- Clase 5Document23 paginiClase 5Biol. Daniela López EscobarÎncă nu există evaluări
- Informe MuscularDocument17 paginiInforme MuscularAlejandro AlvaradoÎncă nu există evaluări
- Tema 10. Fisiología Del MúsculoDocument16 paginiTema 10. Fisiología Del MúsculoKarla GarritsenÎncă nu există evaluări
- La Excitación y Contracción Del Músculo LisoDocument5 paginiLa Excitación y Contracción Del Músculo LisoorutraodnamraÎncă nu există evaluări
- Tejido Muscular ÚltDocument49 paginiTejido Muscular Últvideos MiosÎncă nu există evaluări
- Tejido MuscularDocument4 paginiTejido MuscularJoyce ZambranoÎncă nu există evaluări
- Cuestionario Unidad VI Músculo LisoDocument3 paginiCuestionario Unidad VI Músculo LisoA.D.P.GÎncă nu există evaluări
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesDe la EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesEvaluare: 5 din 5 stele5/5 (1)
- Edema y Signo de GodetDocument15 paginiEdema y Signo de GodetKevin75% (8)
- Modulo 2 UcpqDocument35 paginiModulo 2 UcpqRodrigo De PaulÎncă nu există evaluări
- Glosario de OtorrinolaringologiaDocument6 paginiGlosario de OtorrinolaringologiaStaminados ObaerÎncă nu există evaluări
- Control de La Motilidad GastrointestinalDocument3 paginiControl de La Motilidad GastrointestinalvanessaÎncă nu există evaluări
- Inmunología OcularDocument29 paginiInmunología OcularjramosaliagaÎncă nu există evaluări
- Guia Estudio I Potencial de Membrana y Potencial de AccionDocument9 paginiGuia Estudio I Potencial de Membrana y Potencial de AccionMinastauriel AlassëaÎncă nu există evaluări
- Anomalias Del Sistema InmuneDocument9 paginiAnomalias Del Sistema InmuneGabriela Orozco LopezÎncă nu există evaluări
- Aparato CirculatorioDocument57 paginiAparato CirculatorioMelu IbañezÎncă nu există evaluări
- Aparato Reproductor Masculino y FemeninoDocument4 paginiAparato Reproductor Masculino y FemeninocefuneslpezÎncă nu există evaluări
- Lectura NeurocitologiaDocument7 paginiLectura NeurocitologiaHenryNapaÎncă nu există evaluări
- Glándulas SuprarrenalesDocument33 paginiGlándulas SuprarrenalesAngie SaileÎncă nu există evaluări
- Circulación Fetal :3Document43 paginiCirculación Fetal :3Gabriel Villegas GarciaÎncă nu există evaluări
- Método Anticonceptivo Moco CervicalDocument7 paginiMétodo Anticonceptivo Moco Cervicalcatalina gizziÎncă nu există evaluări
- Nutrición en Animales Invertebrados Y Vertebrados: Clase de Biología-Grado 6° - Colegio Juventudes UnidasDocument18 paginiNutrición en Animales Invertebrados Y Vertebrados: Clase de Biología-Grado 6° - Colegio Juventudes UnidasTatianaFrancoÎncă nu există evaluări
- Fisioterapia en Paralisis FacialDocument36 paginiFisioterapia en Paralisis FacialAurelia IlqÎncă nu există evaluări
- Aparato Reproductor MasculinoDocument11 paginiAparato Reproductor MasculinoNair MartínezÎncă nu există evaluări
- Teoria de La Contraccion MuscularDocument16 paginiTeoria de La Contraccion Musculararmadilloroza100% (1)
- Aparato Reproductor Femenino-MasculinoDocument29 paginiAparato Reproductor Femenino-MasculinoClaudia Elena Guevara HerreraÎncă nu există evaluări
- 2 Anomalias CongenitasDocument78 pagini2 Anomalias CongenitasCLAUDIAÎncă nu există evaluări
- Apuntes de OrtodonciaDocument9 paginiApuntes de OrtodonciaAngelica AmadorÎncă nu există evaluări
- Gliomas AstrocitomasDocument31 paginiGliomas AstrocitomasAaron JaraÎncă nu există evaluări
- La Postura Corporal Equilibrada Consiste en La Alineación Del Cuerpo Con Una Máxima Eficiencia Tanto FisiológicaDocument5 paginiLa Postura Corporal Equilibrada Consiste en La Alineación Del Cuerpo Con Una Máxima Eficiencia Tanto FisiológicaLuciano MartinezÎncă nu există evaluări
- Sistema Genital MasculinoDocument32 paginiSistema Genital MasculinoLopez Alvarado JheisonÎncă nu există evaluări
- Resumen El Veterinario Debe Conocer Las Funciones y Alteraciones Del Sistema CardiovascularDocument14 paginiResumen El Veterinario Debe Conocer Las Funciones y Alteraciones Del Sistema CardiovascularMeyi SanchezÎncă nu există evaluări
- GLOSARIO 1 Farmacognosia y Fitoterapia 1a ParteDocument1 paginăGLOSARIO 1 Farmacognosia y Fitoterapia 1a ParteSophie AvilaÎncă nu există evaluări
- Generalidades de La Mucosa BucalDocument19 paginiGeneralidades de La Mucosa BucalJose Andres Torres0% (2)
- Farmaco MonografiaDocument20 paginiFarmaco MonografiaBruce WilliamÎncă nu există evaluări
- Dia A Dia La Historia de CelesteDocument84 paginiDia A Dia La Historia de CelesteKimberly VargasÎncă nu există evaluări
- Actividad de Mejoramiento de Sistema Endocrino HumanoDocument6 paginiActividad de Mejoramiento de Sistema Endocrino HumanoJose SanchezÎncă nu există evaluări
- Huesos de La CaraDocument17 paginiHuesos de La CaraLaura Fernanda DíazÎncă nu există evaluări