S-ar putea să vă placă și
- Mreznica - 80Document1 paginăMreznica - 80AkicaÎncă nu există evaluări
- Mreznica - 79Document1 paginăMreznica - 79AkicaÎncă nu există evaluări
- Mreznica - 78Document1 paginăMreznica - 78AkicaÎncă nu există evaluări
- Visual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)Document1 paginăVisual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)AkicaÎncă nu există evaluări
- Mreznica - 66Document1 paginăMreznica - 66AkicaÎncă nu există evaluări
- R. M. Shapley and C. Enroth-Cugell: Et AlDocument1 paginăR. M. Shapley and C. Enroth-Cugell: Et AlAkicaÎncă nu există evaluări
- Visual Adaptation and Retinal Gain ControlsDocument1 paginăVisual Adaptation and Retinal Gain ControlsAkicaÎncă nu există evaluări
- Mreznica - 71Document1 paginăMreznica - 71AkicaÎncă nu există evaluări
- Mreznica - 74Document1 paginăMreznica - 74AkicaÎncă nu există evaluări
- Mreznica - 73Document1 paginăMreznica - 73AkicaÎncă nu există evaluări
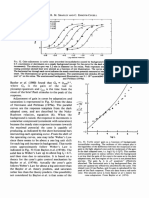
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Document1 paginăEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaÎncă nu există evaluări
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDocument1 pagină6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaÎncă nu există evaluări
- Mreznica - 59Document1 paginăMreznica - 59AkicaÎncă nu există evaluări
- Do Not: 5. Gain Control in PhotoreceptorsDocument1 paginăDo Not: 5. Gain Control in PhotoreceptorsAkicaÎncă nu există evaluări
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDocument1 paginăTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaÎncă nu există evaluări
- Retina - 52Document1 paginăRetina - 52AkicaÎncă nu există evaluări
- Mreznica - 64Document1 paginăMreznica - 64AkicaÎncă nu există evaluări
- 4.3. Horizontal Cells: F I I FDocument1 pagină4.3. Horizontal Cells: F I I FAkicaÎncă nu există evaluări
- Retina - 53 PDFDocument1 paginăRetina - 53 PDFAkicaÎncă nu există evaluări
- Mreznica - 60Document1 paginăMreznica - 60AkicaÎncă nu există evaluări
- Mreznica - 58Document1 paginăMreznica - 58AkicaÎncă nu există evaluări
- Mreznica - 61Document1 paginăMreznica - 61AkicaÎncă nu există evaluări
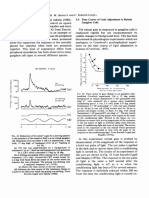
- Time Course of Gain Adjustment in Retinal Ganglion CellsDocument1 paginăTime Course of Gain Adjustment in Retinal Ganglion CellsAkicaÎncă nu există evaluări
- Retina - 52Document1 paginăRetina - 52AkicaÎncă nu există evaluări
- Visual Adaptation and Retinal Gain ControlsDocument1 paginăVisual Adaptation and Retinal Gain ControlsAkicaÎncă nu există evaluări
- Retina - 52Document1 paginăRetina - 52AkicaÎncă nu există evaluări
- Mreznica - 55Document1 paginăMreznica - 55AkicaÎncă nu există evaluări
- Retina - 51Document1 paginăRetina - 51AkicaÎncă nu există evaluări
- 3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsDocument1 pagină3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsAkicaÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Si Pgc5000-En Gen 2 Rev DDocument88 paginiSi Pgc5000-En Gen 2 Rev Dwega deÎncă nu există evaluări
- Anord UL Product RangeDocument8 paginiAnord UL Product RangeSundaresan SabanayagamÎncă nu există evaluări
- Instruction Manual for Lambda EMI Power SupplyDocument66 paginiInstruction Manual for Lambda EMI Power SupplyAntonio Carlos CardosoÎncă nu există evaluări
- Maximum Power Transfer TheoremDocument15 paginiMaximum Power Transfer TheoremAnthony Jay CombateÎncă nu există evaluări
- How To Calculate Peak Data Rate in LTEDocument5 paginiHow To Calculate Peak Data Rate in LTEKashif SaeedÎncă nu există evaluări
- 8.6 MV Feeder Earth-Fault ProtectionDocument20 pagini8.6 MV Feeder Earth-Fault Protectionbiviana81Încă nu există evaluări
- Sharp Lc32bd6x LCD TV SMDocument212 paginiSharp Lc32bd6x LCD TV SMkothuraÎncă nu există evaluări
- MPPTDocument16 paginiMPPTShruheti VaghasiyaÎncă nu există evaluări
- M.SC Thesis PFC - Project - FinalDocument79 paginiM.SC Thesis PFC - Project - FinalRia AlexÎncă nu există evaluări
- My Matlab Code For Random Filter UwbDocument4 paginiMy Matlab Code For Random Filter UwbFawaz SaeedÎncă nu există evaluări
- Chapter 4 Counters PDFDocument37 paginiChapter 4 Counters PDFVeyronie Buggati SteveÎncă nu există evaluări
- Versamate Cleanroom Receptacle PR19Document1 paginăVersamate Cleanroom Receptacle PR19Anonymous llBSa7Încă nu există evaluări
- Dbtp104gui enDocument2 paginiDbtp104gui enMundakir_GNIÎncă nu există evaluări
- BS-200 Service Manual (v1.1)Document123 paginiBS-200 Service Manual (v1.1)Byron Cuevas Mendoza100% (5)
- PM2000 Overview Feature For CustomerDocument11 paginiPM2000 Overview Feature For CustomerChen YujiaÎncă nu există evaluări
- Op-Amp Imperfections: Autozeroing, Correlated Double Sampling, and Chopper StabilizationDocument31 paginiOp-Amp Imperfections: Autozeroing, Correlated Double Sampling, and Chopper StabilizationAndres MauricioÎncă nu există evaluări
- LM2594 0.5 A, Step-Down Switching Regulator: Marking DiagramsDocument27 paginiLM2594 0.5 A, Step-Down Switching Regulator: Marking DiagramsDandy PrasetyoÎncă nu există evaluări
- Testing Electronic ComponentsDocument98 paginiTesting Electronic Componentshajasoftware100% (3)
- GPCB Late Fees CircularDocument2 paginiGPCB Late Fees CircularChirag JainÎncă nu există evaluări
- Hsdpa & Hsupa Challenges For UE Power Amplifier Design: Application NoteDocument50 paginiHsdpa & Hsupa Challenges For UE Power Amplifier Design: Application NoteRamarolahy Dadan'i EliaÎncă nu există evaluări
- Zanussi Electrolux VD54-55 13220950 13220951Document80 paginiZanussi Electrolux VD54-55 13220950 13220951Sam GalaxyÎncă nu există evaluări
- Lecture12 BJT Transistor CircuitsDocument14 paginiLecture12 BJT Transistor CircuitsKartika MunirÎncă nu există evaluări
- Learning Element 3Document17 paginiLearning Element 3niel lunaÎncă nu există evaluări
- Model Yia Absorption Chillers: Renewal PartsDocument12 paginiModel Yia Absorption Chillers: Renewal PartsArshad MahmoodÎncă nu există evaluări
- Problem Set 5Document2 paginiProblem Set 5Saied Aly SalamahÎncă nu există evaluări
- GTP - LT AB Cable - CSPDCLDocument6 paginiGTP - LT AB Cable - CSPDCLNavya cablesÎncă nu există evaluări
- APC Symmetra PX-2Document8 paginiAPC Symmetra PX-2drastir_777Încă nu există evaluări
- B 1815Document4 paginiB 1815Angelo MachadoÎncă nu există evaluări
- MaxiCharger DC Fast-0927Document4 paginiMaxiCharger DC Fast-0927shahrialifizaniÎncă nu există evaluări
- AC#9-Setting Report - Intel - 20.10.20 - R0Document25 paginiAC#9-Setting Report - Intel - 20.10.20 - R0VũDuyTânÎncă nu există evaluări