S-ar putea să vă placă și
- Filogenia y Clasificación de Los AnurosDocument12 paginiFilogenia y Clasificación de Los AnurosJuan PalaciosÎncă nu există evaluări
- Familia CebidaeDocument10 paginiFamilia CebidaeJürgen Bedoya BrandacherÎncă nu există evaluări
- CervidaeDocument3 paginiCervidaeRomel BarrientosÎncă nu există evaluări
- BelgicaDocument10 paginiBelgicaMonica ColimbaÎncă nu există evaluări
- Características Del GrupoDocument6 paginiCaracterísticas Del GrupoGianfranco Edsson Urday YucraÎncă nu există evaluări
- Bufo BufoDocument3 paginiBufo BufoMarco BÎncă nu există evaluări
- Historia Del JabaliDocument5 paginiHistoria Del JabaliMarK RamosÎncă nu există evaluări
- Ficha CIERVO ROJO - Cervus ElaphusDocument2 paginiFicha CIERVO ROJO - Cervus ElaphusMaria RuizÎncă nu există evaluări
- Tema 4 DIVERSIDAD Y CLASIFICACIÓN DE ANFIBIOSDocument6 paginiTema 4 DIVERSIDAD Y CLASIFICACIÓN DE ANFIBIOSsabelacordeiroÎncă nu există evaluări
- El Escuerzo ComúnDocument3 paginiEl Escuerzo ComúnMatias RamosÎncă nu există evaluări
- Mono Araña ComunDocument4 paginiMono Araña ComunAndree SaavedraÎncă nu există evaluări
- Los PelicanosDocument2 paginiLos PelicanosRomina ElizabethÎncă nu există evaluări
- UntitledsjqjsDocument3 paginiUntitledsjqjsAbel ExpósitoÎncă nu există evaluări
- 1er Avance IND. MORFODocument10 pagini1er Avance IND. MORFOariadna ballesterosÎncă nu există evaluări
- FaisánDocument12 paginiFaisánSara AllendesÎncă nu există evaluări
- UrsidaeDocument4 paginiUrsidaeViviana HenaoÎncă nu există evaluări
- UMA Borrego Muflón FINALDocument14 paginiUMA Borrego Muflón FINALTony JMÎncă nu există evaluări
- El Faisan ChinoDocument3 paginiEl Faisan ChinoBorja González-CaminoÎncă nu există evaluări
- Bufo BufoDocument7 paginiBufo BufoJASSSÎncă nu există evaluări
- Negron EspeculadoDocument3 paginiNegron EspeculadoCurriquitoÎncă nu există evaluări
- Trabajo Practico de Diversidad BiologicaDocument6 paginiTrabajo Practico de Diversidad BiologicaFernando QuispeÎncă nu există evaluări
- Bivalvos de Rio Margaritifera AuriculariaDocument13 paginiBivalvos de Rio Margaritifera AuriculariaMarianÎncă nu există evaluări
- El SapoDocument4 paginiEl SapoHENRY ANTONIO FUENTES ESPINOZAÎncă nu există evaluări
- Cria Domino 1Document7 paginiCria Domino 1AlexisÎncă nu există evaluări
- JabaliesDocument6 paginiJabaliesCris AndradeÎncă nu există evaluări
- MarsupialDocument9 paginiMarsupialJuan00000Încă nu există evaluări
- Cerceta AliazulDocument2 paginiCerceta AliazulCurriquitoÎncă nu există evaluări
- Informe CamaleonDocument5 paginiInforme CamaleonJoseÎncă nu există evaluări
- Anade AzulonDocument4 paginiAnade AzulonCurriquitoÎncă nu există evaluări
- La Mosca Del Caballo y Del Venado (Tabanidae)Document0 paginiLa Mosca Del Caballo y Del Venado (Tabanidae)Jorge HdezÎncă nu există evaluări
- Melon CilloDocument2 paginiMelon CilloJose FÎncă nu există evaluări
- Los Opiliones Esos Pequeos DesconocidosDocument3 paginiLos Opiliones Esos Pequeos DesconocidosYani RoibalÎncă nu există evaluări
- Taxonomía Sistemática y Caract de Ballenas en El Golfo de MexDocument3 paginiTaxonomía Sistemática y Caract de Ballenas en El Golfo de MexArline SanchezÎncă nu există evaluări
- Kas AnimalesDocument7 paginiKas AnimalesKassandraMontalvoVeraÎncă nu există evaluări
- Bufo BufoDocument12 paginiBufo BufopinponÎncă nu există evaluări
- Peces y Sus TaxonomiaDocument10 paginiPeces y Sus TaxonomiaDelia MirandaÎncă nu există evaluări
- Informe Final BernasconiDocument17 paginiInforme Final BernasconiandresÎncă nu există evaluări
- Monografia DidelphimorfiaDocument23 paginiMonografia DidelphimorfiaJenny Elizabeth Jimenez GavilanesÎncă nu există evaluări
- ReptilesDocument4 paginiReptilescarlasilva1540Încă nu există evaluări
- Familia Nymphalidae LibroDocument10 paginiFamilia Nymphalidae LibroRuth TerronesÎncă nu există evaluări
- AjoloteDocument8 paginiAjoloteCzar MArtinÎncă nu există evaluări
- CamaleónDocument8 paginiCamaleónHector J. Betancourt LaraÎncă nu există evaluări
- Album de Recursos HidrobiologicosDocument18 paginiAlbum de Recursos Hidrobiologicosjoseluishuanca1224Încă nu există evaluări
- 200000000123Document7 pagini200000000123popijaerworuweoÎncă nu există evaluări
- Sturnus VulgarisDocument22 paginiSturnus VulgarisPaúl NúñezÎncă nu există evaluări
- UrsidaeDocument11 paginiUrsidaejazmin yoselin jara carhuaricraÎncă nu există evaluări
- Phoenicopterus ChilensisDocument3 paginiPhoenicopterus ChilensisHarold RuizÎncă nu există evaluări
- Pato Cordillerano: Reino: Filo: Clase: Orden: Familia: GeneroDocument6 paginiPato Cordillerano: Reino: Filo: Clase: Orden: Familia: GeneroLuis Fernando AguirreÎncă nu există evaluări
- UrsidaeDocument9 paginiUrsidaeGustavo Flores AguireÎncă nu există evaluări
- Flaconiformes - Ob. AvesDocument28 paginiFlaconiformes - Ob. AvesValery ParadoÎncă nu există evaluări
- Tremarctos OrnatusDocument7 paginiTremarctos OrnatusBruno RementeriaÎncă nu există evaluări
- Abecedario de AnimalesDocument33 paginiAbecedario de AnimalesHector MujicaÎncă nu există evaluări
- Micho de Cerro (Modo de Compatibilidad)Document16 paginiMicho de Cerro (Modo de Compatibilidad)Melquiades CastilloÎncă nu există evaluări
- Ursidae - OsoDocument9 paginiUrsidae - OsoJDCLÎncă nu există evaluări
- Familia Saurópsidos Escamosos Especies África Sáhara América AnolesDocument4 paginiFamilia Saurópsidos Escamosos Especies África Sáhara América AnolesFergÎncă nu există evaluări
- AlcedinesDocument5 paginiAlcedinesRuben's OscarÎncă nu există evaluări
- AnfibiosDocument13 paginiAnfibiosGersongenio Rodriguez AÎncă nu există evaluări
- PDF GrullaDocument6 paginiPDF GrullapilarspringÎncă nu există evaluări
- LoroDocument4 paginiLoroVaronil999Încă nu există evaluări
- Manual SP V2Document23 paginiManual SP V2corianoÎncă nu există evaluări
- AcerSystem Guía Del UsuarioDocument27 paginiAcerSystem Guía Del UsuariocorianoÎncă nu există evaluări
- SIMCIMacotelaDocument7 paginiSIMCIMacotelaEmmanuel SalernoÎncă nu există evaluări
- Stairville Led Par 56Document44 paginiStairville Led Par 56corianoÎncă nu există evaluări
- Manual Portatil AcerDocument91 paginiManual Portatil AcerAngelo BiagiÎncă nu există evaluări
- Safety Guide - ES - v2Document7 paginiSafety Guide - ES - v2corianoÎncă nu există evaluări
- Acer Aspire SW3-013Document26 paginiAcer Aspire SW3-013tatanitoÎncă nu există evaluări
- Acer Windows Tablet Safety Guide - ES - v2Document34 paginiAcer Windows Tablet Safety Guide - ES - v2corianoÎncă nu există evaluări
- Ap15aDocument4 paginiAp15acorianoÎncă nu există evaluări
- Acer Regulatory Information and Safety Guide - ES - v4Document31 paginiAcer Regulatory Information and Safety Guide - ES - v4zeuz wildeÎncă nu există evaluări
- MagPi Mini Spanish 01Document20 paginiMagPi Mini Spanish 01Carlos López Brage100% (2)
- Certificación AP15A3RDocument4 paginiCertificación AP15A3RcorianoÎncă nu există evaluări
- Empezar Raspberry PiDocument24 paginiEmpezar Raspberry PiFreddy LarenasÎncă nu există evaluări
- PDFDocument29 paginiPDFAntonio FloresÎncă nu există evaluări
- Prosign 130Document68 paginiProsign 130corianoÎncă nu există evaluări
- Como Obtener Roseicollis LutinosDocument2 paginiComo Obtener Roseicollis LutinoscorianoÎncă nu există evaluări
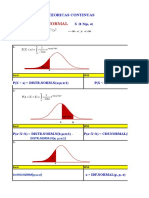
- TRABAJO #3 Distribución Normal EstandarDocument2 paginiTRABAJO #3 Distribución Normal EstandarDavid Panca Humpiri0% (1)
- Familia Compuesta TesisDocument86 paginiFamilia Compuesta Tesiskellyn luqueÎncă nu există evaluări
- PC1 FamiliaySucesionesDocument17 paginiPC1 FamiliaySucesionesLuis David ChafloqueÎncă nu există evaluări
- Antropologia AvanzadaDocument4 paginiAntropologia AvanzadaRaúl Armando Santana RivasÎncă nu există evaluări
- Capítulo 3 EnanosDocument57 paginiCapítulo 3 EnanosAndres RomeroÎncă nu există evaluări
- Salutacion en El Dia Del Padre PetroleroDocument1 paginăSalutacion en El Dia Del Padre PetroleroMaría Del Carmen Hidalgo AguilarÎncă nu există evaluări
- INCAUSADOSRAFINADocument5 paginiINCAUSADOSRAFINAalexabarredaÎncă nu există evaluări
- Fuentes de La InformaciónDocument3 paginiFuentes de La InformaciónRolando Jesus Flores BelizarioÎncă nu există evaluări
- Derecho de Familia - Soledad Quintana VillarDocument12 paginiDerecho de Familia - Soledad Quintana VillarClaudiaÎncă nu există evaluări
- Herramientas de IntervenciónDocument81 paginiHerramientas de IntervenciónVIVIANA LILLO RAMÍREZÎncă nu există evaluări
- Distribuciones Contínuas 2018 - 000Document47 paginiDistribuciones Contínuas 2018 - 000Moises Juarez0% (1)
- Ampliacion de Demanda Dentro Del Proceso CivilDocument2 paginiAmpliacion de Demanda Dentro Del Proceso Civilgilmar zacariasÎncă nu există evaluări
- 01 - Ahora Soy Yo La ProtaDocument142 pagini01 - Ahora Soy Yo La ProtaOriana EspinozaÎncă nu există evaluări
- Ejercicios Distribucion Normal EstandarDocument4 paginiEjercicios Distribucion Normal EstandarHumberto muñoz bravoÎncă nu există evaluări
- Acerca de La AperturaDocument12 paginiAcerca de La AperturaAnabelÎncă nu există evaluări
- Sexualidad, Riesgo U OportunidadDocument2 paginiSexualidad, Riesgo U OportunidadHEIDY ISABEL100% (2)
- Vocabulary Ingles Preuniversitario 1Document7 paginiVocabulary Ingles Preuniversitario 1Reyes Farias IsraelÎncă nu există evaluări
- GreciaDocument2 paginiGreciaKaren VillalobosÎncă nu există evaluări
- The Letter To The Magi - Carta A Los Reyes MagosDocument4 paginiThe Letter To The Magi - Carta A Los Reyes MagospabolecÎncă nu există evaluări
- ConvivenciaDocument2 paginiConvivenciaAnonymous LvOeimHRÎncă nu există evaluări
- ADOPCIÓNDocument24 paginiADOPCIÓNDíaz Luis100% (1)
- Biografia LenguaDocument679 paginiBiografia LenguaArkanaÎncă nu există evaluări
- Derecho Romano Tema 3Document15 paginiDerecho Romano Tema 3Alberto Zamora100% (1)
- H. Auto Final Declarando Con Lugar Diligencias de Adopción.Document5 paginiH. Auto Final Declarando Con Lugar Diligencias de Adopción.Luis SaputÎncă nu există evaluări
- COTRINA - Resumen de Lounsbury (1964)Document1 paginăCOTRINA - Resumen de Lounsbury (1964)Felipe AlvaradoÎncă nu există evaluări
- El ConcubinatoDocument18 paginiEl ConcubinatoMillan Ana MercedesÎncă nu există evaluări
- Sucesión NotarialDocument9 paginiSucesión Notarialaldemar sanchezÎncă nu există evaluări
- Espanhol 4Document4 paginiEspanhol 4Carlos EduardoÎncă nu există evaluări
- Diccionario TicoDocument10 paginiDiccionario TicoJuan Bosco Bustos SalazarÎncă nu există evaluări
- Botánica. Angiospermas. Clase MagnoliopsidaDocument49 paginiBotánica. Angiospermas. Clase Magnoliopsidamarijoya100% (1)