S-ar putea să vă placă și
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967De la EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterÎncă nu există evaluări
- NT 5mmbleed 1Document6 paginiNT 5mmbleed 1Diana MihaiÎncă nu există evaluări
- Interaction of Some Local Anaesthetics and Antisickling Drugs With Calcium-Carrier Proteins in BiomenbranesDocument8 paginiInteraction of Some Local Anaesthetics and Antisickling Drugs With Calcium-Carrier Proteins in BiomenbranesClement BewajiÎncă nu există evaluări
- Comparative Uptake Studies of Bioadhesive and Non-Bioadhesive Nanoparticles in Human Intestinal Cell Lines and Rats: The Effect of Mucus On Particle Adsorption and TransportDocument2 paginiComparative Uptake Studies of Bioadhesive and Non-Bioadhesive Nanoparticles in Human Intestinal Cell Lines and Rats: The Effect of Mucus On Particle Adsorption and TransportBrenda OrtizÎncă nu există evaluări
- Nae Et Al 2008Document7 paginiNae Et Al 2008BeatriceMihaelaRaduÎncă nu există evaluări
- 2009 AemDocument8 pagini2009 AemAndrew SingerÎncă nu există evaluări
- Aerobacter Aerogenes J. Bacteriol. 1966 Bernofsky 1404 14Document11 paginiAerobacter Aerogenes J. Bacteriol. 1966 Bernofsky 1404 14ClaudioÎncă nu există evaluări
- CapsaicinDocument12 paginiCapsaicinExlonk Gil PeláezÎncă nu există evaluări
- Estudio 2Document5 paginiEstudio 2Gabriel GarridoÎncă nu există evaluări
- Differential Expression ... Unicellular ChlorophyteDocument7 paginiDifferential Expression ... Unicellular ChlorophyteAJITHKUMARÎncă nu există evaluări
- Pyrenomonas Salina: Isolation, Physical Map and Gene Map of Mitochondrial DNA From The CryptomonadDocument8 paginiPyrenomonas Salina: Isolation, Physical Map and Gene Map of Mitochondrial DNA From The CryptomonadJuvelin AuliaÎncă nu există evaluări
- Exacerbation of Acetaminophen Hepatotoxicity by The Anthelmentic Drug FenbendazoleDocument6 paginiExacerbation of Acetaminophen Hepatotoxicity by The Anthelmentic Drug FenbendazoleSkyalesÎncă nu există evaluări
- Orcinol MethodDocument8 paginiOrcinol MethodSHAILEE GAURÎncă nu există evaluări
- 1 FullDocument9 pagini1 FullFauziana NurhanisahÎncă nu există evaluări
- Jamur Tiram 1Document6 paginiJamur Tiram 1Muhammad AkbarÎncă nu există evaluări
- Inhibition of Peptidoglycan Biosynthesis in Gram-Positive Bacteria by LY146032Document7 paginiInhibition of Peptidoglycan Biosynthesis in Gram-Positive Bacteria by LY146032SachithÎncă nu există evaluări
- The Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andDocument9 paginiThe Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andAvishekh SinhaÎncă nu există evaluări
- Magni2004 Article EnzymologyOfNADHomeostasisInMaDocument16 paginiMagni2004 Article EnzymologyOfNADHomeostasisInMaLeidy Constanza Villalobos GonzalezÎncă nu există evaluări
- Recombinant: Human Dnase I Reduces The Viscosity of Cystic Fibrosis SputumDocument5 paginiRecombinant: Human Dnase I Reduces The Viscosity of Cystic Fibrosis SputumGuhan KAÎncă nu există evaluări
- JB 82 4 582-588 1961Document7 paginiJB 82 4 582-588 1961laasyagudiÎncă nu există evaluări
- Sloane1981 - Science - CathepsinsDocument4 paginiSloane1981 - Science - CathepsinsPilar AufrastoÎncă nu există evaluări
- 2006 Cabagna y Col.Document9 pagini2006 Cabagna y Col.Mariana CabagnaÎncă nu există evaluări
- Colloids and Surfaces B: BiointerfacesDocument7 paginiColloids and Surfaces B: BiointerfacesAFRIZAÎncă nu există evaluări
- Glutamine's role in neutrophil superoxide production and NADPH oxidase component expressionDocument6 paginiGlutamine's role in neutrophil superoxide production and NADPH oxidase component expressionAstynnia GanÎncă nu există evaluări
- Communication: Nephrogenic Diabetes InsipidusDocument4 paginiCommunication: Nephrogenic Diabetes Insipiduslinda aprilÎncă nu există evaluări
- Cancer Epidemiol Biomarkers Prev-1999-Hecht-907-13Document8 paginiCancer Epidemiol Biomarkers Prev-1999-Hecht-907-13Ivon Buitrago VillanuevaÎncă nu există evaluări
- PDFDocument11 paginiPDFAstridz Monsteqszia SilvanaÎncă nu există evaluări
- Carotenoids Other Than Vitamin A — III: Third International Symposium on Carotenoids Other Than Vitamin ADe la EverandCarotenoids Other Than Vitamin A — III: Third International Symposium on Carotenoids Other Than Vitamin AÎncă nu există evaluări
- P450-Mediated Carbon–Carbon Ring Closure in Lathyrane Diterpenoid BiosynthesisDocument5 paginiP450-Mediated Carbon–Carbon Ring Closure in Lathyrane Diterpenoid BiosynthesisEdithÎncă nu există evaluări
- J Pharmacol Exp Ther-1999-Fisher-1134-42Document9 paginiJ Pharmacol Exp Ther-1999-Fisher-1134-42krantithesisÎncă nu există evaluări
- Xanthorrhizol Has A Potential To Attenuate The High Dose Cisplatin-Induced Nephrotoxicity in MiceDocument6 paginiXanthorrhizol Has A Potential To Attenuate The High Dose Cisplatin-Induced Nephrotoxicity in MiceFajri RifaldiÎncă nu există evaluări
- Paper 6Document4 paginiPaper 6Harry HartÎncă nu există evaluări
- 365 Full PDFDocument8 pagini365 Full PDFMiguel Machaca Flores (QuimioFarma)Încă nu există evaluări
- In-Vitro Cellular Uptake and Transport Study of 9-NitrocamptothecinDocument10 paginiIn-Vitro Cellular Uptake and Transport Study of 9-NitrocamptothecinMonica TurnerÎncă nu există evaluări
- Carbon Nanotubes As Intracellular Protein Transporters: Generality and Biological FunctionalityDocument6 paginiCarbon Nanotubes As Intracellular Protein Transporters: Generality and Biological Functionality227986Încă nu există evaluări
- HW 0440Document8 paginiHW 0440JaanvhiÎncă nu există evaluări
- Rhodospirillum Rubrum Has A Family I PyrophosphataseDocument4 paginiRhodospirillum Rubrum Has A Family I PyrophosphataseCarolina RicárdezÎncă nu există evaluări
- Literature Review: Expense ofDocument43 paginiLiterature Review: Expense ofAnn DassÎncă nu există evaluări
- Fukamachi Et Al-2008-Cancer ScienceDocument6 paginiFukamachi Et Al-2008-Cancer ScienceMonica Cabezas DipazÎncă nu există evaluări
- Said AhmedDocument19 paginiSaid Ahmedahmed mahdiÎncă nu există evaluări
- Fagan Et Al. 1999 - Quantitation-Of-Oxidative-Damage-To-Tissue-Proteins - 1999 - The-International-Journal-of-Biochemistry-Cell-BiologyDocument7 paginiFagan Et Al. 1999 - Quantitation-Of-Oxidative-Damage-To-Tissue-Proteins - 1999 - The-International-Journal-of-Biochemistry-Cell-BiologyAntonios MatsakasÎncă nu există evaluări
- Quercetin Reduces p21-Ras Expression in Colon CancerDocument8 paginiQuercetin Reduces p21-Ras Expression in Colon CancerBenePicarÎncă nu există evaluări
- Rhodobacter Capsulad: Microbial Photodegradation of Aminoarenes Metabolism of 2-Amino-4-Nitro Henol byDocument12 paginiRhodobacter Capsulad: Microbial Photodegradation of Aminoarenes Metabolism of 2-Amino-4-Nitro Henol byntphquynhÎncă nu există evaluări
- Graphene Oxide in HER-2-positive CancerDocument7 paginiGraphene Oxide in HER-2-positive CancerSoraya Torres GazeÎncă nu există evaluări
- Carcinogen Metabolism: BC DeckerDocument3 paginiCarcinogen Metabolism: BC DeckerTanu WijayaÎncă nu există evaluări
- Pääbo Mitochondrial NAR 1988Document14 paginiPääbo Mitochondrial NAR 1988jaafari mohamedÎncă nu există evaluări
- Chloroplast Import of Four Carotenoid Biosynthetic EnzymesDocument9 paginiChloroplast Import of Four Carotenoid Biosynthetic EnzymesNithin RÎncă nu există evaluări
- Role of Nadph:Cytochrome C Reducã-Aseand Dt-Diaphorase in The Biotransformation of Mitomycin C1Document7 paginiRole of Nadph:Cytochrome C Reducã-Aseand Dt-Diaphorase in The Biotransformation of Mitomycin C1Krys tallÎncă nu există evaluări
- 2006 Cannabidiol-Induced Apoptosis in Human Leukemia Cells (Mol Pharmacol, McKallip)Document12 pagini2006 Cannabidiol-Induced Apoptosis in Human Leukemia Cells (Mol Pharmacol, McKallip)Rune Christian FoynÎncă nu există evaluări
- Gene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialDocument6 paginiGene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialHuong TranÎncă nu există evaluări
- The Flavouring Phytochemical 2-Pentanone Reduces Prostaglandin Production and COX-2 Expression in Colon Cancer CellsDocument4 paginiThe Flavouring Phytochemical 2-Pentanone Reduces Prostaglandin Production and COX-2 Expression in Colon Cancer CellsyosuacahyaÎncă nu există evaluări
- Research ArticleDocument9 paginiResearch ArticleMinh Đức HoàngÎncă nu există evaluări
- Manoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Document8 paginiManoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Collin LoganÎncă nu există evaluări
- CDK2 RoscovitineDocument9 paginiCDK2 RoscovitineputriÎncă nu există evaluări
- Saponins (Effects On Membrane Permeability)Document10 paginiSaponins (Effects On Membrane Permeability)andrewalamÎncă nu există evaluări
- The Warburg Effect and Its Cancer Therapeutic Implications: Mini ReviewDocument8 paginiThe Warburg Effect and Its Cancer Therapeutic Implications: Mini ReviewAndrea RangelÎncă nu există evaluări
- Pan KDocument9 paginiPan Kafrank2Încă nu există evaluări
- Metabolic Engineering of Carotenoid Accumulation in E.coliDocument5 paginiMetabolic Engineering of Carotenoid Accumulation in E.coliAJITHKUMARÎncă nu există evaluări
- NIH Public Access: Author ManuscriptDocument27 paginiNIH Public Access: Author ManuscriptChristian AlvisÎncă nu există evaluări
- Biotransformation of Benzo A Pyrene and Other Polycyclic Aromatic HydrocarbonsDocument18 paginiBiotransformation of Benzo A Pyrene and Other Polycyclic Aromatic HydrocarbonsgeoffreyÎncă nu există evaluări
- T3D0026Document3 paginiT3D0026Micawber7Încă nu există evaluări
- Danish ImperativeDocument3 paginiDanish ImperativeMicawber7Încă nu există evaluări
- Seattle Bellevuepark Barbera EbrochureDocument1 paginăSeattle Bellevuepark Barbera EbrochureMicawber7Încă nu există evaluări
- 05 - An Evaluation of Tetramethylbenzidine As A Presumptive Test For BloodDocument6 pagini05 - An Evaluation of Tetramethylbenzidine As A Presumptive Test For BloodMicawber7Încă nu există evaluări
- Phonological AsymmetriesDocument1 paginăPhonological AsymmetriesMicawber7Încă nu există evaluări
- Amalia E. Gnanadesikan - Markedness and Faithdulness Constraints in Child Phonology PDFDocument48 paginiAmalia E. Gnanadesikan - Markedness and Faithdulness Constraints in Child Phonology PDFMicawber7Încă nu există evaluări
- Adams - Government of Empty Subjects in Factive Clausal ComplementsDocument5 paginiAdams - Government of Empty Subjects in Factive Clausal ComplementsMicawber7Încă nu există evaluări
- Adolfo Ausn - A-Traces and Semantic InterpretationDocument6 paginiAdolfo Ausn - A-Traces and Semantic InterpretationMicawber7Încă nu există evaluări
- Cristina Job Schmitt - Aspect and The Syntax of Noun Phrases PDFDocument223 paginiCristina Job Schmitt - Aspect and The Syntax of Noun Phrases PDFMicawber7Încă nu există evaluări
- Deriving Effects by Crossing ScalesDocument5 paginiDeriving Effects by Crossing ScalesMicawber7Încă nu există evaluări
- Bermudez-Otero - Underlyingly Nonmoraic Coda Consonants, Faithfulness, and Sympathy PDFDocument28 paginiBermudez-Otero - Underlyingly Nonmoraic Coda Consonants, Faithfulness, and Sympathy PDFMicawber7Încă nu există evaluări
- Aissen - Differential Object Marking - Iconicity Vs EconomyDocument6 paginiAissen - Differential Object Marking - Iconicity Vs EconomyMicawber7Încă nu există evaluări
- Revised Schedule of Lab and Discussion Sessions # Date Duratio N Locatio N TopicDocument1 paginăRevised Schedule of Lab and Discussion Sessions # Date Duratio N Locatio N TopicMicawber7Încă nu există evaluări
- Ealize Orph EME: H Pl. H Pl. Pl. H PLDocument7 paginiEalize Orph EME: H Pl. H Pl. Pl. H PLMicawber7Încă nu există evaluări
- Case Studies PDFDocument6 paginiCase Studies PDFMicawber7Încă nu există evaluări
- Smith 2001 PDFDocument12 paginiSmith 2001 PDFMicawber7Încă nu există evaluări
- Hazardous Substance Fact Sheet: Right To KnowDocument6 paginiHazardous Substance Fact Sheet: Right To KnowMicawber7Încă nu există evaluări
- Rhodamine DyesDocument5 paginiRhodamine DyesMicawber7Încă nu există evaluări
- TP 62Document2 paginiTP 62Micawber7Încă nu există evaluări
- Anttila - Morphologically Conditioned Phonological AlternationsDocument44 paginiAnttila - Morphologically Conditioned Phonological AlternationsMicawber7Încă nu există evaluări
- Aissen - Differential Object Marking - Iconicity Vs Economy PDFDocument44 paginiAissen - Differential Object Marking - Iconicity Vs Economy PDFMicawber7Încă nu există evaluări
- tp62 PDFDocument242 paginitp62 PDFMicawber7Încă nu există evaluări
- Benzidine MetabolismDocument4 paginiBenzidine MetabolismMicawber7Încă nu există evaluări
- Community Acquired PneumoniaDocument63 paginiCommunity Acquired PneumoniaLet BorlagdanÎncă nu există evaluări
- Biochemistry For Anesthesiologists and IntensivistsDocument11 paginiBiochemistry For Anesthesiologists and IntensivistsAndi tenri sanda datuÎncă nu există evaluări
- PR Menarini PDX Ab RBCDocument2 paginiPR Menarini PDX Ab RBCvyasakandarpÎncă nu există evaluări
- Eucalyptus The Genus Eucalyptus PDFDocument433 paginiEucalyptus The Genus Eucalyptus PDFPeter Wilhelmsen100% (5)
- 1413272524world10 15 14 PDFDocument38 pagini1413272524world10 15 14 PDFCoolerAdsÎncă nu există evaluări
- Jobs Safety Analysis for Radiography WorkDocument4 paginiJobs Safety Analysis for Radiography WorkGan KusumaÎncă nu există evaluări
- Respiratory Physiology AnswersDocument4 paginiRespiratory Physiology AnswersRamya100% (2)
- WHO Recommended Test For TBDocument40 paginiWHO Recommended Test For TBJasminÎncă nu există evaluări
- Voice Production in Singing and SpeakingBased On Scientific Principles (Fourth Edition, Revised and Enlarged) by Mills, Wesley, 1847-1915Document165 paginiVoice Production in Singing and SpeakingBased On Scientific Principles (Fourth Edition, Revised and Enlarged) by Mills, Wesley, 1847-1915Gutenberg.org100% (3)
- Narrative Report On School Nursing - EditedDocument16 paginiNarrative Report On School Nursing - EditedAbby_Cacho_9151100% (2)
- 1 PBDocument21 pagini1 PBDewi Puspita SariÎncă nu există evaluări
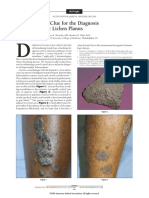
- A Novel Visual Clue For The Diagnosis of Hypertrophic Lichen PlanusDocument1 paginăA Novel Visual Clue For The Diagnosis of Hypertrophic Lichen Planus600WPMPOÎncă nu există evaluări
- Bls - Fbao - First AidDocument172 paginiBls - Fbao - First AidMaria Regina Castro Gabriel100% (1)
- Personal health data for nursing studentDocument8 paginiPersonal health data for nursing studentJenny AjocÎncă nu există evaluări
- Atp4 - 25x13 Casuality Evacuation PDFDocument88 paginiAtp4 - 25x13 Casuality Evacuation PDFbdavid2012-1Încă nu există evaluări
- AlgorithmACLS Tachycardia 200612Document1 paginăAlgorithmACLS Tachycardia 200612YassarÎncă nu există evaluări
- Pedigree Analysis: (Cf. Chapters 4.4, 5.2, 6.2 of Textbook)Document11 paginiPedigree Analysis: (Cf. Chapters 4.4, 5.2, 6.2 of Textbook)Kristyne OliciaÎncă nu există evaluări
- Seminar Molar DistalizationDocument130 paginiSeminar Molar DistalizationMuhsina EyyanathilÎncă nu există evaluări
- Parallax Xray For Maxillary CanineDocument3 paginiParallax Xray For Maxillary Canineabdul samad noorÎncă nu există evaluări
- Tilapia 4Document69 paginiTilapia 4Annisa MeilaniÎncă nu există evaluări
- CRRT Continuous Renal Replacement Therapies ImplementationDocument28 paginiCRRT Continuous Renal Replacement Therapies ImplementationyamtotlÎncă nu există evaluări
- A Client With A Brain Tumor: Nursing Care PlanDocument1 paginăA Client With A Brain Tumor: Nursing Care Planshabatat2002Încă nu există evaluări
- Tender No 38 S Haad PT 2014Document26 paginiTender No 38 S Haad PT 2014Sundar Ramasamy100% (1)
- Tracheostomy Care With ChecklistDocument5 paginiTracheostomy Care With ChecklistHollan GaliciaÎncă nu există evaluări
- JMDH 400734 Application Benefits and Limitations of Telepharmacy For PDocument9 paginiJMDH 400734 Application Benefits and Limitations of Telepharmacy For PIrma RahmawatiÎncă nu există evaluări
- Indian ListenerDocument103 paginiIndian ListenerTareq Aziz100% (1)
- Vocal Cord ParalysisDocument74 paginiVocal Cord ParalysisWaqas KhanÎncă nu există evaluări
- Abdominal UltrasoundDocument6 paginiAbdominal Ultrasounds0800841739100% (1)
- Jubilant Life Sciences Receives ANDA Approval For Rosuvastatin Calcium Tablets (Company Update)Document3 paginiJubilant Life Sciences Receives ANDA Approval For Rosuvastatin Calcium Tablets (Company Update)Shyam SunderÎncă nu există evaluări
- Lab Test Results 7 2015Document3 paginiLab Test Results 7 2015Antzee Dancee0% (1)