S-ar putea să vă placă și
- Dam Lecture 14 - Design Criteria Earth DamsDocument4 paginiDam Lecture 14 - Design Criteria Earth DamsCarolineMwitaMoseregaÎncă nu există evaluări
- SSN Melaka SMK Seri Kota 2021 Annual Training Plan: Athletes Name Training ObjectivesDocument2 paginiSSN Melaka SMK Seri Kota 2021 Annual Training Plan: Athletes Name Training Objectivessiapa kahÎncă nu există evaluări
- Micro RNADocument35 paginiMicro RNAkritichd100% (1)
- CH 10 Gene ExpressionDocument23 paginiCH 10 Gene ExpressionerichaasÎncă nu există evaluări
- February 2019Document4 paginiFebruary 2019sagar manghwaniÎncă nu există evaluări
- Creamy and Thick Mushroom Soup: IngredientsDocument8 paginiCreamy and Thick Mushroom Soup: IngredientsSheila Mae AramanÎncă nu există evaluări
- Cash and Cash Equivalents ReviewerDocument4 paginiCash and Cash Equivalents ReviewerEileithyia KijimaÎncă nu există evaluări
- Gene SilencingDocument18 paginiGene SilencingNAMRA RASHEEDÎncă nu există evaluări
- B65a RRH2x40-4R UHGC SPDocument71 paginiB65a RRH2x40-4R UHGC SPNicolás RuedaÎncă nu există evaluări
- Benefits of Silicone Coated Glass InsulatorsDocument10 paginiBenefits of Silicone Coated Glass InsulatorsMuhammad Waseem AhmadÎncă nu există evaluări
- Transcription and RNA ProcessingDocument38 paginiTranscription and RNA ProcessingRishi Kumar100% (1)
- Capstone-ModDocument25 paginiCapstone-ModMohammad Ryyan PumbagulÎncă nu există evaluări
- 20MSRBT028 GeneSilencingDocument29 pagini20MSRBT028 GeneSilencing200409120010Încă nu există evaluări
- Gene Silencing: Presented By: Parvez ShaikhDocument29 paginiGene Silencing: Presented By: Parvez ShaikhsankarÎncă nu există evaluări
- Gene SilencingDocument29 paginiGene SilencingDaud KhanÎncă nu există evaluări
- Gene SileningDocument30 paginiGene SileningSajjad AhmadÎncă nu există evaluări
- 2019 BTEC 5360 ClassDocument31 pagini2019 BTEC 5360 ClassSeayoung LEEÎncă nu există evaluări
- RNA InterferenceDocument26 paginiRNA InterferenceAjayChandrakarÎncă nu există evaluări
- Aaryan Gupta 2018B1A70775HDocument7 paginiAaryan Gupta 2018B1A70775HAaryan GuptaÎncă nu există evaluări
- Gene SilencingDocument7 paginiGene SilencingMaliha JahanÎncă nu există evaluări
- MICRORNAs: A REVIEW STUDYDocument10 paginiMICRORNAs: A REVIEW STUDYIOSR Journal of PharmacyÎncă nu există evaluări
- Antisense OligoDocument40 paginiAntisense Oligoinigo100% (1)
- RNAi MphilDocument28 paginiRNAi Mphilrameeshasaleem01Încă nu există evaluări
- Gene Silencing - Antisense RNA & SiRNADocument18 paginiGene Silencing - Antisense RNA & SiRNAvenkatsuriyaprakash100% (1)
- Mechanism & FunctionsDocument7 paginiMechanism & FunctionsShaher Bano MirzaÎncă nu există evaluări
- Rna InterferenceDocument19 paginiRna InterferencePROFESSORÎncă nu există evaluări
- Gene Silencing: Presented byDocument23 paginiGene Silencing: Presented byRouf Ahmad100% (1)
- Non-Coding RNADocument2 paginiNon-Coding RNAvanderbroÎncă nu există evaluări
- RNAi NotesDocument6 paginiRNAi Notesaman jaiswalÎncă nu există evaluări
- Rna InterferenceDocument10 paginiRna InterferencewaktoleÎncă nu există evaluări
- Post-Transcriptional Gene Silencing: Sanjeev Sharma, Aarti Bhardwaj, Shalini Jain and Hariom YadavDocument28 paginiPost-Transcriptional Gene Silencing: Sanjeev Sharma, Aarti Bhardwaj, Shalini Jain and Hariom YadavGhayas Muhammad KhanÎncă nu există evaluări
- Gene Silencing Gene Silencing: Niranjana.S.Karandikar Msc-I Roll No.12Document21 paginiGene Silencing Gene Silencing: Niranjana.S.Karandikar Msc-I Roll No.12Niranjana KarandikarÎncă nu există evaluări
- Gene Silencing: How Does It Works?Document3 paginiGene Silencing: How Does It Works?Joon Bok NamleeÎncă nu există evaluări
- Antisense RNA: Example: The Flavr Savr TomatoDocument10 paginiAntisense RNA: Example: The Flavr Savr Tomatomamut343Încă nu există evaluări
- Sirna: Hort Nterfering Ibo Ucleic CidDocument16 paginiSirna: Hort Nterfering Ibo Ucleic CidSimon HollÎncă nu există evaluări
- RNA Interference: Navigation SearchDocument13 paginiRNA Interference: Navigation Searchrathiak2Încă nu există evaluări
- Lecture 4 AntisenseDocument48 paginiLecture 4 AntisensesankarÎncă nu există evaluări
- RNA Interference Class NotesDocument4 paginiRNA Interference Class NotesmanaleepandyaÎncă nu există evaluări
- Sirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardDocument7 paginiSirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardChristian Moises CasasÎncă nu există evaluări
- Modifications Controlling RNADocument5 paginiModifications Controlling RNADiego RodriguezÎncă nu există evaluări
- Jackson Review Microrna 2007Document13 paginiJackson Review Microrna 2007iuventasÎncă nu există evaluări
- Lecture 16 Regulating Genes by RNADocument19 paginiLecture 16 Regulating Genes by RNAzhy20040213Încă nu există evaluări
- Gene Silencing TechniquesDocument15 paginiGene Silencing TechniquesShaher Bano Mirza100% (2)
- Regulation of Gene ExpressionDocument7 paginiRegulation of Gene ExpressionGodwin UnimashiÎncă nu există evaluări
- Elbashir Et Al. 2001Document12 paginiElbashir Et Al. 2001Benedikt EngelÎncă nu există evaluări
- Chapter 8 LodishDocument52 paginiChapter 8 LodishablucÎncă nu există evaluări
- Micro Rna and EpigDocument12 paginiMicro Rna and EpigNayágara MoreiraÎncă nu există evaluări
- Mi RNAworldDocument10 paginiMi RNAworldMARIA DE JESUS SANCHEZ MURGUIAÎncă nu există evaluări
- RNA Interference: Hannon, Nature 418:244-251Document24 paginiRNA Interference: Hannon, Nature 418:244-251EdwinsMusicÎncă nu există evaluări
- Life and Times of An Rna L3Document4 paginiLife and Times of An Rna L3katieÎncă nu există evaluări
- Application of Short Hairpin RNAs (shRNAs) To StudyDocument6 paginiApplication of Short Hairpin RNAs (shRNAs) To StudyBhaskar RoyÎncă nu există evaluări
- CellbioDocument13 paginiCellbiojafameg801Încă nu există evaluări
- Rna Interference FinalDocument29 paginiRna Interference FinalParvathi BabuÎncă nu există evaluări
- Background: Rna Interference: The Next Genetics Revolution?Document4 paginiBackground: Rna Interference: The Next Genetics Revolution?Shiv KumarÎncă nu există evaluări
- The Use of Artifi Cial MicroRNA Technology To Control Gene Expression in Arabidopsis ThalianaDocument14 paginiThe Use of Artifi Cial MicroRNA Technology To Control Gene Expression in Arabidopsis ThalianaThắng Trần BảoÎncă nu există evaluări
- RNA Interference: An Approach For Sequence-Specific Knockdown of MrnaDocument49 paginiRNA Interference: An Approach For Sequence-Specific Knockdown of MrnasungroÎncă nu există evaluări
- Transcription WorkingDocument51 paginiTranscription Workingapi-3858544Încă nu există evaluări
- Introduction To Proteomics and Genomics: Asma Asif FA17-BSI-002 DR IbrahimDocument3 paginiIntroduction To Proteomics and Genomics: Asma Asif FA17-BSI-002 DR IbrahimMuhammad Faizan AsifÎncă nu există evaluări
- 2013-Diversifying MicroRNA Sequence and FunctionDocument14 pagini2013-Diversifying MicroRNA Sequence and FunctionJorge Hantar Touma LazoÎncă nu există evaluări
- The Circulating Transcriptome As A Source of Cancer Liquid Biopsy BiomarkersDocument32 paginiThe Circulating Transcriptome As A Source of Cancer Liquid Biopsy BiomarkersRyan MalolotÎncă nu există evaluări
- Gene ExpressionDocument44 paginiGene ExpressionCahya MaharaniÎncă nu există evaluări
- Gene ExpressionDocument44 paginiGene Expression20.020 Cahya MaharaniÎncă nu există evaluări
- RNA Cleavage Products Generated by Antisense OligoDocument13 paginiRNA Cleavage Products Generated by Antisense OligoHimanshu PanchalÎncă nu există evaluări
- RNA Function Prediction: Yongsheng Li, Juan Xu, Tingting Shao, Yunpeng Zhang, Hong Chen, and Xia LiDocument12 paginiRNA Function Prediction: Yongsheng Li, Juan Xu, Tingting Shao, Yunpeng Zhang, Hong Chen, and Xia Lireda rashwanÎncă nu există evaluări
- Transgene SLIENCINGDocument35 paginiTransgene SLIENCINGRajaram Krishnasamy100% (1)
- RNA Interference: Hannon, Nature 418:244-251Document24 paginiRNA Interference: Hannon, Nature 418:244-251VaishaliSharmaÎncă nu există evaluări
- Régulation de L'éxpréssion Génétique (1) - CopieDocument163 paginiRégulation de L'éxpréssion Génétique (1) - CopieWahuba RahmaniÎncă nu există evaluări
- Rahu Yantra Kal Sarp Yantra: Our RecommendationsDocument2 paginiRahu Yantra Kal Sarp Yantra: Our RecommendationsAbhijeet DeshmukkhÎncă nu există evaluări
- Edgie A. Tenerife BSHM 1108: Page 1 of 4Document4 paginiEdgie A. Tenerife BSHM 1108: Page 1 of 4Edgie TenerifeÎncă nu există evaluări
- RB Boiler Product SpecsDocument4 paginiRB Boiler Product Specsachmad_silmiÎncă nu există evaluări
- SAT Subject Chemistry SummaryDocument25 paginiSAT Subject Chemistry SummaryYoonho LeeÎncă nu există evaluări
- Compensation ManagementDocument2 paginiCompensation Managementshreekumar_scdlÎncă nu există evaluări
- Presentation of DR Rai On Sahasrara Day Medical SessionDocument31 paginiPresentation of DR Rai On Sahasrara Day Medical SessionRahul TikkuÎncă nu există evaluări
- SET 2022 Gstr1Document1 paginăSET 2022 Gstr1birpal singhÎncă nu există evaluări
- TS802 - Support StandardDocument68 paginiTS802 - Support StandardCassy AbulenciaÎncă nu există evaluări
- Aliant Ommunications: VCL-2709, IEEE C37.94 To E1 ConverterDocument2 paginiAliant Ommunications: VCL-2709, IEEE C37.94 To E1 ConverterConstantin UdreaÎncă nu există evaluări
- BlackCat ManualDocument14 paginiBlackCat ManualNestor Marquez-DiazÎncă nu există evaluări
- Weekly Meal Prep GuideDocument7 paginiWeekly Meal Prep Guideandrew.johnson3112Încă nu există evaluări
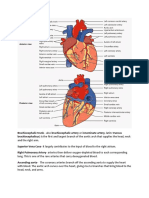
- Brachiocephalic TrunkDocument3 paginiBrachiocephalic TrunkstephÎncă nu există evaluări
- II092 - Horiz & Vert ULSs With Serial InputsDocument4 paginiII092 - Horiz & Vert ULSs With Serial InputsJibjab7Încă nu există evaluări
- Microbial Communities From Arid Environments On A Global Scale. A Systematic ReviewDocument12 paginiMicrobial Communities From Arid Environments On A Global Scale. A Systematic ReviewAnnaÎncă nu există evaluări
- Grundfos Data Booklet MMSrewindablesubmersiblemotorsandaccessoriesDocument52 paginiGrundfos Data Booklet MMSrewindablesubmersiblemotorsandaccessoriesRashida MajeedÎncă nu există evaluări
- Cough PDFDocument3 paginiCough PDFKASIA SyÎncă nu există evaluări
- Tomography: Tomography Is Imaging by Sections or Sectioning Through The Use of AnyDocument6 paginiTomography: Tomography Is Imaging by Sections or Sectioning Through The Use of AnyJames FranklinÎncă nu există evaluări
- Consent CertificateDocument5 paginiConsent Certificatedhanu2399Încă nu există evaluări
- 4th Summative Science 6Document2 pagini4th Summative Science 6brian blase dumosdosÎncă nu există evaluări
- Latihan Soal Bahasa Inggris 2Document34 paginiLatihan Soal Bahasa Inggris 2Anita KusumastutiÎncă nu există evaluări
- Soal 2-3ADocument5 paginiSoal 2-3Atrinanda ajiÎncă nu există evaluări
- 1 BSC Iriigation Engineering 2018 19 Std1Document70 pagini1 BSC Iriigation Engineering 2018 19 Std1Kwasi BempongÎncă nu există evaluări