S-ar putea să vă placă și
- Marcadores CardíacosDocument9 paginiMarcadores CardíacosAngel AguirreÎncă nu există evaluări
- Músculos Del TóraxDocument4 paginiMúsculos Del TóraxLeslie LedesmaÎncă nu există evaluări
- Procesos Carnicos - Modulo-Julio2012Document179 paginiProcesos Carnicos - Modulo-Julio2012Carlos Mauricio Sanchez83% (6)
- Resumen Guyton y Hall CAPITULO 81Document11 paginiResumen Guyton y Hall CAPITULO 81Daniel Jaramillo AponteÎncă nu există evaluări
- Embriologia Sistema OsteomuscularDocument26 paginiEmbriologia Sistema OsteomuscularKristell Quezada100% (1)
- 4.8 Contracción Muscular PDFDocument6 pagini4.8 Contracción Muscular PDFErick Peñaloza100% (1)
- Histologia Usmp 2019 I - Tejido EpitelialDocument57 paginiHistologia Usmp 2019 I - Tejido EpitelialMilagros Blas Pizarro0% (1)
- Quimica Clinica en El LaboratorioDocument51 paginiQuimica Clinica en El LaboratorioLeydi Yudith Angarita Bautista100% (12)
- Pared Anterolateral PDFDocument8 paginiPared Anterolateral PDFerikaalexandrahÎncă nu există evaluări
- Histo Resumen Ojo y OidoDocument10 paginiHisto Resumen Ojo y OidoNOHELY JOLI AYALA YABARÎncă nu există evaluări
- Fisiologia Veterinaria Cap3 Contraccion MuscularDocument13 paginiFisiologia Veterinaria Cap3 Contraccion MuscularAlejandroz GonzalezÎncă nu există evaluări
- Seminario de Tejido MuscularDocument19 paginiSeminario de Tejido Musculardanly castilloÎncă nu există evaluări
- Unidad MotoraDocument18 paginiUnidad MotoraMédecin Adrian TGÎncă nu există evaluări
- 7.2) El Cromosoma - Organizacion Del ADNDocument45 pagini7.2) El Cromosoma - Organizacion Del ADNSzzczzÎncă nu există evaluări
- Estructura Del EspermatozoideDocument7 paginiEstructura Del EspermatozoideedispaÎncă nu există evaluări
- Origen Embriológico de RiñonDocument1 paginăOrigen Embriológico de RiñonGlenn GarcíaÎncă nu există evaluări
- Generalidades Del Sistema OsteomioarticularDocument7 paginiGeneralidades Del Sistema OsteomioarticularPatricioViquez0% (1)
- 26 Abdomen. GeneralidadesDocument17 pagini26 Abdomen. GeneralidadesFranklin garryÎncă nu există evaluări
- CorazónDocument10 paginiCorazónJL Díaz0% (1)
- Tejido Oseo y Cartilaginoso PDFDocument63 paginiTejido Oseo y Cartilaginoso PDFAna RomeroÎncă nu există evaluări
- Tejido MuscularDocument36 paginiTejido MuscularGiovanniMonsalvoÎncă nu există evaluări
- Capitulo 55 FisioDocument2 paginiCapitulo 55 FisioVickyFingadoÎncă nu există evaluări
- ApoptosisDocument17 paginiApoptosisLuis CarlosÎncă nu există evaluări
- El EosinofiloDocument4 paginiEl Eosinofilopepe islasÎncă nu există evaluări
- Histología RiñónDocument7 paginiHistología RiñónSneider Alexander Garcia TascoÎncă nu există evaluări
- Sinapsis NeuromuscularDocument4 paginiSinapsis NeuromuscularFabio SotoÎncă nu există evaluări
- Unidad 7 Sistema DigestivoDocument35 paginiUnidad 7 Sistema DigestivoGerardo RodriguezÎncă nu există evaluări
- La Formación de Mórula, Blastocisto y GastrulaciónDocument21 paginiLa Formación de Mórula, Blastocisto y GastrulaciónMarlene ToledanoÎncă nu există evaluări
- Embriologia Del Sistema CardiovascularDocument13 paginiEmbriologia Del Sistema CardiovascularMirgelys VargasÎncă nu există evaluări
- Resumen Pagina 422 - 423 Higado (Gartner)Document4 paginiResumen Pagina 422 - 423 Higado (Gartner)Edwin Chávez HuamánÎncă nu există evaluări
- Cuestionario Del Aparato Reproductor FemeninoDocument3 paginiCuestionario Del Aparato Reproductor FemeninoLeon LÎncă nu există evaluări
- Tejido CartilaginosoDocument100 paginiTejido Cartilaginosojessica moscosoÎncă nu există evaluări
- Trabajo Del Tejido Epitelial....Document4 paginiTrabajo Del Tejido Epitelial....Maria Castro Cruz0% (1)
- Histología de La Artrosis de RodillaDocument9 paginiHistología de La Artrosis de RodillaAntonio VazquezÎncă nu există evaluări
- Histología Del Sistema Reproductor MasculinoDocument48 paginiHistología Del Sistema Reproductor MasculinoSofiajrfÎncă nu există evaluări
- VÍAS URINARIAS DR - Nixor 2019Document13 paginiVÍAS URINARIAS DR - Nixor 2019Carol Jennifer Núñez Campos100% (1)
- Informe Anatomía Humana II Sesión 3Document7 paginiInforme Anatomía Humana II Sesión 3Angie Flores MamaniÎncă nu există evaluări
- CesarDocument10 paginiCesarapi-3699557100% (1)
- Atlas HistologíaDocument20 paginiAtlas HistologíaMarlÎncă nu există evaluări
- Tejido CartilaginosoDocument36 paginiTejido CartilaginosoJessi Ramirez UÎncă nu există evaluări
- Prevencion de La Coagulacion Sanguinea en El Sistema Vascular NormalDocument2 paginiPrevencion de La Coagulacion Sanguinea en El Sistema Vascular NormalBryan Ty Castro GonzalesÎncă nu există evaluări
- Cuestionario 9Document3 paginiCuestionario 9diegosteman100% (2)
- Trastorno Del Crecimiento y Desarrollo CelularDocument3 paginiTrastorno Del Crecimiento y Desarrollo CelularRodrigo Ochoa CalleÎncă nu există evaluări
- Generalidades Del Sistema OsteomuscularDocument37 paginiGeneralidades Del Sistema Osteomuscularjuan bruzualÎncă nu există evaluări
- Actividad 16 AnatomíaDocument10 paginiActividad 16 AnatomíaNicole QuinchuelaÎncă nu există evaluări
- Vertebras y ToraxDocument4 paginiVertebras y ToraxRochu GiordanoÎncă nu există evaluări
- Fisiologia Muscular Upsjb 4ºclaseDocument40 paginiFisiologia Muscular Upsjb 4ºclaseKrystel GonzalesÎncă nu există evaluări
- Cuáles Son Las Partes de La SarcomeraDocument2 paginiCuáles Son Las Partes de La Sarcomeraruben dario puello royalÎncă nu există evaluări
- Desarrollo PlacentarioDocument12 paginiDesarrollo PlacentarioAngel Vincenzo CorbinoÎncă nu există evaluări
- Anatomía Fisiológica Del Músculo EsqueléticoDocument7 paginiAnatomía Fisiológica Del Músculo EsqueléticoBárbara Garcés Jarpa100% (1)
- APUNTES EmbriologiaDocument5 paginiAPUNTES Embriologiagabriel ChuquimiaÎncă nu există evaluări
- Cerebro Configuración InternaDocument10 paginiCerebro Configuración InternaNino' Peace50% (2)
- Embriologia Sistema Renal Urogenital 14Document37 paginiEmbriologia Sistema Renal Urogenital 14AnaAguerrevereÎncă nu există evaluări
- Desarrollo Del Tubo Intestinal, Páncreas y DiafragmaDocument6 paginiDesarrollo Del Tubo Intestinal, Páncreas y Diafragmaeliana bustamante0% (1)
- Tejido Linfático y Órganos Linfáticos MapaDocument10 paginiTejido Linfático y Órganos Linfáticos MapaSara OlveraÎncă nu există evaluări
- Histología Del OídoDocument11 paginiHistología Del OídoluisÎncă nu există evaluări
- Informe 3 Embriología SeminarioDocument8 paginiInforme 3 Embriología SeminarioEmery Sofía C. D. BustamanteÎncă nu există evaluări
- Embriologia - Esqueleto AxialDocument8 paginiEmbriologia - Esqueleto AxialMajency SilvaÎncă nu există evaluări
- Tejido EpitelialDocument44 paginiTejido EpitelialHenry Petit RodriguezÎncă nu există evaluări
- Seminario N 5Document16 paginiSeminario N 5Nina Saboya CórdovaÎncă nu există evaluări
- Grupo 11 Tejido Hematopoyético PreguntasDocument2 paginiGrupo 11 Tejido Hematopoyético PreguntasMateoo06Încă nu există evaluări
- Guia - Enzimas PDFDocument8 paginiGuia - Enzimas PDFAdrián GpÎncă nu există evaluări
- Contracción Del Músculo Esquelético, Excitación y Contracción Del Musculo Liso.Document66 paginiContracción Del Músculo Esquelético, Excitación y Contracción Del Musculo Liso.Louis ViceÎncă nu există evaluări
- Musculo LisoDocument12 paginiMusculo LisoAlex Rodriguez Casas100% (1)
- Sistema MuscularDocument70 paginiSistema MuscularJeremy Baker Jacobo Vásquez100% (1)
- Clase Adhesión y Matriz Celular 2019 JSDocument94 paginiClase Adhesión y Matriz Celular 2019 JSmacaÎncă nu există evaluări
- Núcleo CelularDocument71 paginiNúcleo CelularmacaÎncă nu există evaluări
- Clase Doc FertisexDocument3 paginiClase Doc FertisexmacaÎncă nu există evaluări
- Morfología Del Desarrollo Del Aparato GenitalDocument7 paginiMorfología Del Desarrollo Del Aparato GenitalmacaÎncă nu există evaluări
- CitogeneticaDocument71 paginiCitogeneticamacaÎncă nu există evaluări
- Hierro Mitocondrial en La Ataxia de FriedreichDocument7 paginiHierro Mitocondrial en La Ataxia de FriedreichmacaÎncă nu există evaluări
- Apuntes Biofisica (Musculo Esqueletico)Document27 paginiApuntes Biofisica (Musculo Esqueletico)Camila NievesÎncă nu există evaluări
- Diagnostico Con Enzimas Cardiacas. Manejo Infarto Agudo MiocardioDocument8 paginiDiagnostico Con Enzimas Cardiacas. Manejo Infarto Agudo MiocardioSara OchoaÎncă nu există evaluări
- Tema 23 - Oposición EFDocument36 paginiTema 23 - Oposición EFVíctor Sánchez AguilarÎncă nu există evaluări
- Quimica Clinica en El LaboratorioDocument51 paginiQuimica Clinica en El LaboratorioMinet GaticaÎncă nu există evaluări
- Enzimas CardíacasDocument2 paginiEnzimas CardíacasCarlos SegovianoÎncă nu există evaluări
- Biofisica 2021Document51 paginiBiofisica 2021Mel LantesÎncă nu există evaluări
- Trauma CardiacoDocument67 paginiTrauma CardiacoRosalieÎncă nu există evaluări
- 5 MuscularDocument49 pagini5 MuscularMaacaBodoCanalesÎncă nu există evaluări
- Repaso Tema 2Document6 paginiRepaso Tema 2Sebastian GonzalezÎncă nu există evaluări
- 1 m1 U1Document44 pagini1 m1 U1Anonymous 8ZfVHt100% (1)
- Hipertermia MalignaDocument26 paginiHipertermia Malignaapi-36995570% (1)
- Fisiologia MuscularDocument24 paginiFisiologia MuscularJean CuencaÎncă nu există evaluări
- Enzimas - 18DDocument37 paginiEnzimas - 18DJohan ZÎncă nu există evaluări
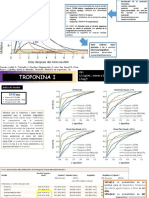
- Troponina I Exposicicon Curva Roc ...Document5 paginiTroponina I Exposicicon Curva Roc ...Ruth QGÎncă nu există evaluări
- Anatomia II Primer ParcialDocument9 paginiAnatomia II Primer Parcialbladimir100% (1)
- Ultraestructura y Bioquímica Del Músculo EsqueléticoDocument33 paginiUltraestructura y Bioquímica Del Músculo EsqueléticomaseÎncă nu există evaluări
- Generalidades de MiologíaDocument65 paginiGeneralidades de MiologíaRoger GarciaÎncă nu există evaluări
- 5.3-Postquirurgico Cardio-FinalDocument17 pagini5.3-Postquirurgico Cardio-FinaldarwinÎncă nu există evaluări
- Historieta. Contracción MuscularDocument5 paginiHistorieta. Contracción MuscularCarlos MarguzÎncă nu există evaluări
- Clase 1 Gastr - Introduccion Al Musculo LisoDocument17 paginiClase 1 Gastr - Introduccion Al Musculo LisoFelipe Ignacio Gutierrez BarriosÎncă nu există evaluări
- Carne y CárnicosDocument8 paginiCarne y CárnicosAndrés Flores SilesÎncă nu există evaluări
- Teoría de La Refrigeración y CongelaciónDocument27 paginiTeoría de La Refrigeración y CongelaciónAnonymous jowdOKkW850% (2)
- Apunte Completo de Fisiologia Del EjercicioDocument83 paginiApunte Completo de Fisiologia Del EjercicioFede uXxÎncă nu există evaluări