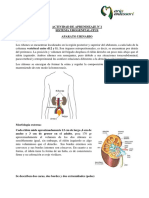
S-ar putea să vă placă și
- Obtencion de AcetilenoDocument16 paginiObtencion de AcetilenoZhaarlee Aurelijus89% (28)
- LABORATORIO DE BIOQUIMICA - Reconocimiento y Propiedades de Los LípidosDocument33 paginiLABORATORIO DE BIOQUIMICA - Reconocimiento y Propiedades de Los LípidosJENNY ALEXANDRA ROSAS VARGASÎncă nu există evaluări
- Factores AbioticosDocument13 paginiFactores AbioticosIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- Covid PeruDocument1 paginăCovid PeruIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- Charlas de Seguridad de 5Document14 paginiCharlas de Seguridad de 5Ibeth Mónica Machaca CatacoraÎncă nu există evaluări
- Charlas de Seguridad de 5Document14 paginiCharlas de Seguridad de 5Ibeth Mónica Machaca CatacoraÎncă nu există evaluări
- Minera AntapiteDocument4 paginiMinera AntapiteJulio Saraeen Bautista CondoriÎncă nu există evaluări
- Leyes de Seguridad IntroducciónDocument1 paginăLeyes de Seguridad IntroducciónIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- EXTRACCIÓNDocument3 paginiEXTRACCIÓNIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- EiaDocument1 paginăEiaIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- MinamDocument5 paginiMinamIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- La CombustiónDocument1 paginăLa CombustiónIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- EiaDocument1 paginăEiaIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- EIA Cerro VerdeDocument1 paginăEIA Cerro VerdeIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- MinamDocument5 paginiMinamIbeth Mónica Machaca CatacoraÎncă nu există evaluări
- Documento Lesiones Periodontales AgudasDocument9 paginiDocumento Lesiones Periodontales AgudasMaria isabel MosquraÎncă nu există evaluări
- SEMANA 2 BioenergeticaDocument31 paginiSEMANA 2 BioenergeticaKeiko Kimberly RamosÎncă nu există evaluări
- SeramDocument23 paginiSeram980Denis FernandezÎncă nu există evaluări
- Partes Del OJODocument9 paginiPartes Del OJOElizabeth GarcíaÎncă nu există evaluări
- Actividad de Aprendizaje N°1 Fisiopatologia FarmaciaDocument11 paginiActividad de Aprendizaje N°1 Fisiopatologia FarmaciaRosa Maria Pillco CcahuaÎncă nu există evaluări
- EIA Mineria PDFDocument132 paginiEIA Mineria PDFDante Eduardo Mormontoy RuelasÎncă nu există evaluări
- Enfermedad Renal AteroembólicaDocument7 paginiEnfermedad Renal AteroembólicaKarla Ruiz RobledoÎncă nu există evaluări
- Datos Estudiantes-20Document12 paginiDatos Estudiantes-20JhoelElvisVivasMancha0% (1)
- Homenaje Biológico A Mickey MouseDocument9 paginiHomenaje Biológico A Mickey MouseDonde Busquemos Obra Teatral50% (2)
- Resumen de La Teoría de DarwinDocument4 paginiResumen de La Teoría de DarwinCecy GuerraÎncă nu există evaluări
- Treybalr Ejercicios CoutiñoDocument16 paginiTreybalr Ejercicios CoutiñoVargas Soriano MaRcela50% (2)
- Resumen de La Odontologia ForenseDocument24 paginiResumen de La Odontologia ForenseLic. Ivan Orlando Ramírez SantanaÎncă nu există evaluări
- Secme-9274 1Document74 paginiSecme-9274 1RICARDOÎncă nu există evaluări
- Cuestionario Tema 1Document6 paginiCuestionario Tema 1gary moralesÎncă nu există evaluări
- Control de La Calidad de La LecheDocument10 paginiControl de La Calidad de La LecheElmer Ramos QuispeÎncă nu există evaluări
- Mediastino PosteriorDocument6 paginiMediastino PosteriorRafaelÎncă nu există evaluări
- MalariaDocument14 paginiMalariaGaby NicoleÎncă nu există evaluări
- Kappa Caseína TuestaDocument16 paginiKappa Caseína TuestaERWIN F. TUESTA HZO.Încă nu există evaluări
- Tema 9 Soluciones Intravenosas Seccion 04Document34 paginiTema 9 Soluciones Intravenosas Seccion 04Josebeth RisquezÎncă nu există evaluări
- ANA PatronesDocument28 paginiANA PatronespedroÎncă nu există evaluări
- INFGENETICA YwbdpeditadoDocument19 paginiINFGENETICA YwbdpeditadoEliana Michelle Parra Barajas100% (1)
- Driving Pressure and Mechanical Power - New Targets For VILI Prevention 2017 (3) Ingles - It.es PDFDocument10 paginiDriving Pressure and Mechanical Power - New Targets For VILI Prevention 2017 (3) Ingles - It.es PDFmaraca12345Încă nu există evaluări
- 9648-Texto Del Artículo-9555-1-10-20220723Document30 pagini9648-Texto Del Artículo-9555-1-10-20220723Yolanda TamayoÎncă nu există evaluări
- Riesgos Psicosociales de Enfermería QuirúrgicaDocument3 paginiRiesgos Psicosociales de Enfermería QuirúrgicaProfa ConsueloÎncă nu există evaluări
- Guía Didáctica UP1Document4 paginiGuía Didáctica UP1tiziana.batistuttiÎncă nu există evaluări
- Curvas de Absorcion MaizDocument16 paginiCurvas de Absorcion MaizHarvin GudielÎncă nu există evaluări
- Salud Publica y EpidemiologiaDocument5 paginiSalud Publica y EpidemiologiaRocío América Pereira MonsalveÎncă nu există evaluări
- Fisiopatologia Gastrointestinal IIDocument107 paginiFisiopatologia Gastrointestinal IIEva Nina PorcoÎncă nu există evaluări
- NoradrenalinaDocument6 paginiNoradrenalinaLuis José UrbanoÎncă nu există evaluări