S-ar putea să vă placă și
- Schiestl & Johnson 2013 Pollinator Mediate Evolution Floral SignalsDocument9 paginiSchiestl & Johnson 2013 Pollinator Mediate Evolution Floral SignalsMaria Del Carmen Hernandez ValenciaÎncă nu există evaluări
- Koski 2020 the Role of Sensory Drive in FloralDocument13 paginiKoski 2020 the Role of Sensory Drive in FloralAziz Ait AkkaÎncă nu există evaluări
- Senses and Signals Review Current ScienceDocument10 paginiSenses and Signals Review Current ScienceNiranjana Prasanna VenkateswaranÎncă nu există evaluări
- Floral Morphs of Justicia Adhatoda L. Differ in Fruit and Seed, But Not Oral, Traits or Pollinator VisitationDocument14 paginiFloral Morphs of Justicia Adhatoda L. Differ in Fruit and Seed, But Not Oral, Traits or Pollinator VisitationEapsa BerryÎncă nu există evaluări
- Akter 2017 Effects of Small Scale Clustering oDocument14 paginiAkter 2017 Effects of Small Scale Clustering oAziz Ait AkkaÎncă nu există evaluări
- Electron Microscopic Studies of Some Anticancer Plants Pollen GrainsDocument8 paginiElectron Microscopic Studies of Some Anticancer Plants Pollen GrainsIJAR JOURNALÎncă nu există evaluări
- A Global Test of The Pollination Syndrome HypothesisDocument10 paginiA Global Test of The Pollination Syndrome HypothesisConstanza Andía AvendañoÎncă nu există evaluări
- Variation in Floral Fragrance Compounds of Epidendrum ciliare OrchidDocument7 paginiVariation in Floral Fragrance Compounds of Epidendrum ciliare OrchidValeria TavaresÎncă nu există evaluări
- Flower Reproduction in PlantsDocument22 paginiFlower Reproduction in Plantstechnically whoÎncă nu există evaluări
- New Phytologist - 2021 - Thorogood - Endoparasitic Plants and Fungi Show Evolutionary Convergence Across PhylogeneticDocument9 paginiNew Phytologist - 2021 - Thorogood - Endoparasitic Plants and Fungi Show Evolutionary Convergence Across PhylogeneticDan Mora GarcíaÎncă nu există evaluări
- 1 s2.0 S0169534705001928 MainDocument8 pagini1 s2.0 S0169534705001928 Mainankush prakashÎncă nu există evaluări
- Sexual Reproduction in Flowering Plants: A Fascinating OrganDocument23 paginiSexual Reproduction in Flowering Plants: A Fascinating OrganPrathyusha MannemÎncă nu există evaluări
- Odorant ReceptorsDocument7 paginiOdorant ReceptorschanwilonxÎncă nu există evaluări
- Microbial fingerprints reveal plant-pollinator interactionsDocument7 paginiMicrobial fingerprints reveal plant-pollinator interactionsJorgeVictorMauriceLiraÎncă nu există evaluări
- Neu Big 2011Document30 paginiNeu Big 2011Alexander DamiánÎncă nu există evaluări
- Leonard Et Al-2011-Functional EcologyDocument9 paginiLeonard Et Al-2011-Functional EcologyElisa FuentesÎncă nu există evaluări
- Sexual Reproduction in Flowering Plants: 2.1 F - A F O ADocument23 paginiSexual Reproduction in Flowering Plants: 2.1 F - A F O AVikram SkÎncă nu există evaluări
- Plant Kingdom Botany E-Book for Public ExamsDe la EverandPlant Kingdom Botany E-Book for Public ExamsÎncă nu există evaluări
- Bogarin - 2016 - Orquidea EpidendrumDocument34 paginiBogarin - 2016 - Orquidea EpidendrumDavid AngaritaÎncă nu există evaluări
- Table Rocks Curriculum: Pollination Partners ObjectiveDocument10 paginiTable Rocks Curriculum: Pollination Partners ObjectiveMAXIMO AROCUTIPA CATACORAÎncă nu există evaluări
- Bohs, 2001 - Revision of Solanum Section CyphomandropsisDocument86 paginiBohs, 2001 - Revision of Solanum Section CyphomandropsisCristian Chico VinuezaÎncă nu există evaluări
- genes-13-00678Document14 paginigenes-13-00678muhammad dailamiÎncă nu există evaluări
- Schiestl - 2010 - The Evolution of Floral Scent and Insect Chemical Communication PDFDocument14 paginiSchiestl - 2010 - The Evolution of Floral Scent and Insect Chemical Communication PDFMaria Camila Garcia GiraldoÎncă nu există evaluări
- Jdarago2002 - Trichromatic Color Vision in PrimatesDocument6 paginiJdarago2002 - Trichromatic Color Vision in PrimatesAnkanÎncă nu există evaluări
- 2023 Plants LamiumDocument17 pagini2023 Plants LamiumProf. Dr. Wafaa AmerÎncă nu există evaluări
- Within-Individual Phenotypic Plasticity in Owers Fosters Pollination Niche ShiftDocument12 paginiWithin-Individual Phenotypic Plasticity in Owers Fosters Pollination Niche ShiftMichelle CañizaresÎncă nu există evaluări
- Pollen Morphological Studies of Selected Plants of Asteraceae FamilyDocument8 paginiPollen Morphological Studies of Selected Plants of Asteraceae FamilyIJRASETPublicationsÎncă nu există evaluări
- Zygomorphic Flowers Have Fewer Potential Pollinator SpeciesDocument6 paginiZygomorphic Flowers Have Fewer Potential Pollinator SpeciesDaniel Toro MezaÎncă nu există evaluări
- Agr 516 - Laboratory ManualDocument20 paginiAgr 516 - Laboratory ManualAiman WafiyÎncă nu există evaluări
- Adua SeminarDocument16 paginiAdua SeminarIBRAHIM ISMAIL ADUAÎncă nu există evaluări
- BENITEZ-VIEYRA Et Al-2009-Journal of Evolutionary BiologyDocument9 paginiBENITEZ-VIEYRA Et Al-2009-Journal of Evolutionary BiologyAlfredo PerettiÎncă nu există evaluări
- Rationalised 2023-24Document25 paginiRationalised 2023-24Archana shuklaÎncă nu există evaluări
- Biology Pollination Lab Spring 09 - FinalDocument9 paginiBiology Pollination Lab Spring 09 - FinalRafal WhittingtonÎncă nu există evaluări
- Cardoso-Gustavson Et Al. (2018) Unidirectional Transitions in Nectar Gain and Loss Suggest Food Deception Is A Stable Evolutionary Strategy in EpidendrumDocument13 paginiCardoso-Gustavson Et Al. (2018) Unidirectional Transitions in Nectar Gain and Loss Suggest Food Deception Is A Stable Evolutionary Strategy in EpidendrumJuan AndréÎncă nu există evaluări
- Biology Pollination Updated 2017Document11 paginiBiology Pollination Updated 2017Rafal WhittingtonÎncă nu există evaluări
- Jove 89 52083Document8 paginiJove 89 52083Hassan WajidÎncă nu există evaluări
- Articulo en Ingles D. MelanogasterDocument6 paginiArticulo en Ingles D. Melanogasterzafiro TorresÎncă nu există evaluări
- Flowering Plants ExplainedDocument4 paginiFlowering Plants ExplainedGodfrey ObingoÎncă nu există evaluări
- Foods: Spectroscopic Discrimination of Bee Pollen by Composition, Color, and Botanical OriginDocument13 paginiFoods: Spectroscopic Discrimination of Bee Pollen by Composition, Color, and Botanical OriginvmsolartecÎncă nu există evaluări
- Fruend 2011 Ecology LettDocument9 paginiFruend 2011 Ecology LettAngel Eduardo Moreno AlboresÎncă nu există evaluări
- Lesson Plan: Plants & InsectsDocument9 paginiLesson Plan: Plants & InsectsGemerlyn VallesÎncă nu există evaluări
- Plant Morphology - WikipediaDocument57 paginiPlant Morphology - WikipediaBashiir NuurÎncă nu există evaluări
- Neural analysis of sound frequency in insectsDocument9 paginiNeural analysis of sound frequency in insectsLê ĐứcÎncă nu există evaluări
- Laboratory Activity No.3Document1 paginăLaboratory Activity No.3Elaine MoralesÎncă nu există evaluări
- Trends Plant Sci 2006 BrennerDocument7 paginiTrends Plant Sci 2006 BrennerAnijuali91Încă nu există evaluări
- 304-Article Text-998-1-10-20100302 PDFDocument5 pagini304-Article Text-998-1-10-20100302 PDFristonsitorusÎncă nu există evaluări
- Haddad 2008Document9 paginiHaddad 2008Gabriela Bonilla PerassoÎncă nu există evaluări
- Lab Report 1 Pollination Lab - Elise Grossman - GradedDocument8 paginiLab Report 1 Pollination Lab - Elise Grossman - GradedLisaa GrossmanÎncă nu există evaluări
- Coevolution of Species in Mutualistic RelationshipsDocument3 paginiCoevolution of Species in Mutualistic RelationshipsMudassara HussainÎncă nu există evaluări
- Bimodal Pollination Systems in Andean MelastomataceaeDocument14 paginiBimodal Pollination Systems in Andean MelastomataceaeCamilo Barrios CardozoÎncă nu există evaluări
- Red Leaves, Insects and Coevolution: A Red Herring?: H. Martin Schaefer and David M. WilkinsonDocument3 paginiRed Leaves, Insects and Coevolution: A Red Herring?: H. Martin Schaefer and David M. Wilkinsonsri anika cahayuÎncă nu există evaluări
- ecs2.2943Document15 paginiecs2.2943Aziz Ait AkkaÎncă nu există evaluări
- Ecologia Evolutiva 1Document15 paginiEcologia Evolutiva 1JESSICA HERRERA SANCHEZÎncă nu există evaluări
- Ecology 7Document16 paginiEcology 7JESSICA HERRERA SANCHEZÎncă nu există evaluări
- Group 1 - Flowers' Secret Signal To Bees and Other Amazing Nanotechnologies Hidden in PlantsDocument4 paginiGroup 1 - Flowers' Secret Signal To Bees and Other Amazing Nanotechnologies Hidden in PlantsChâu Anh TrầnÎncă nu există evaluări
- Pollinators and Its Behavior On Delonix Regia (Boj. Ex Hook) Raf. (Family: Caesalpiniaceae)Document8 paginiPollinators and Its Behavior On Delonix Regia (Boj. Ex Hook) Raf. (Family: Caesalpiniaceae)IOSRjournalÎncă nu există evaluări
- Why We Believe in Creation Not EvolutionDocument242 paginiWhy We Believe in Creation Not EvolutionJoeÎncă nu există evaluări
- Ansci 30 Slaughter HouseDocument34 paginiAnsci 30 Slaughter HouseDieanne MaeÎncă nu există evaluări
- Rood S ApproachDocument33 paginiRood S Approachmedway physio teamÎncă nu există evaluări
- Honey Bee Complaint As Community Policing For Scouting Against PCA Act, 1960: Restoring 5 FreedomsDocument31 paginiHoney Bee Complaint As Community Policing For Scouting Against PCA Act, 1960: Restoring 5 FreedomsNaresh KadyanÎncă nu există evaluări
- R-Controlled VowelsDocument10 paginiR-Controlled VowelsShelby BruzzeseÎncă nu există evaluări
- Selalu Kurang Dari 6 Menit (Satu Dari 10 Tikus Memiliki 8Document9 paginiSelalu Kurang Dari 6 Menit (Satu Dari 10 Tikus Memiliki 8Annisa Dwi YantiÎncă nu există evaluări
- Grammar Exercises - Simple Past TenseDocument6 paginiGrammar Exercises - Simple Past TensePatty JonasÎncă nu există evaluări
- Main Idea 2Document3 paginiMain Idea 2api-234576762100% (3)
- INAPHDocument4 paginiINAPHSri VasuÎncă nu există evaluări
- Concept Map - Abby !Document2 paginiConcept Map - Abby !Abegail Abaygar100% (3)
- Comparative Animal Physiology: A Guide to Thermoregulation and MetabolismDocument7 paginiComparative Animal Physiology: A Guide to Thermoregulation and MetabolismWwwanand111Încă nu există evaluări
- Crocheted Uterus PatternDocument2 paginiCrocheted Uterus Patternweasley123100% (4)
- Cousins Cum 4 1Document15 paginiCousins Cum 4 1ty.mcgillÎncă nu există evaluări
- Office Head Titles MunicipalityDocument2 paginiOffice Head Titles MunicipalityGeo AngeloÎncă nu există evaluări
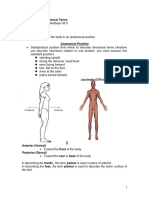
- Anatomical TermsDocument8 paginiAnatomical TermsDAGUMAN, FIONA DEI L.Încă nu există evaluări
- Mage The Awakening - Intruders - Encounters With The Abyss PDFDocument228 paginiMage The Awakening - Intruders - Encounters With The Abyss PDFJohnRichardWuethrich100% (2)
- Darwin Comes To TownDocument7 paginiDarwin Comes To TownTadeoÎncă nu există evaluări
- Phonetic Animals ListDocument6 paginiPhonetic Animals ListJOHN. J MESAÎncă nu există evaluări
- Noun Practice PagesDocument6 paginiNoun Practice PagesMaggie Plby100% (1)
- The Illustrated Horse S Foot PDFDocument273 paginiThe Illustrated Horse S Foot PDFGIANELLA ESPINOZA MENDOZA100% (5)
- Neurons Stop Motion AnimationDocument2 paginiNeurons Stop Motion Animationapi-495006167Încă nu există evaluări
- I. Multiple Choice. Choose The Letter of The Best AnswerDocument3 paginiI. Multiple Choice. Choose The Letter of The Best AnswerKeannoÎncă nu există evaluări
- Kids 1 2015 FinalDocument3 paginiKids 1 2015 FinalSilvina HillarÎncă nu există evaluări
- Ethical Guidelines PsychologyDocument3 paginiEthical Guidelines PsychologyManal_99xoÎncă nu există evaluări
- A Naive Bride ExcellentDocument52 paginiA Naive Bride ExcellentAhmed BilalÎncă nu există evaluări
- Microbiological Monitoring - RODACDocument2 paginiMicrobiological Monitoring - RODACdeden.drj19Încă nu există evaluări
- Lounging Leafer Giraffe DollDocument33 paginiLounging Leafer Giraffe DollDana Richards100% (1)
- Types of ClaimsDocument14 paginiTypes of ClaimsCatherine Joy ManaloÎncă nu există evaluări
- Task 6 and 7 ANIMALSDocument2 paginiTask 6 and 7 ANIMALSRonny AlexanderÎncă nu există evaluări
- ProverbDocument38 paginiProverbAnkita GoelÎncă nu există evaluări