S-ar putea să vă placă și
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- 14 PDFDocument21 pagini14 PDFPalanisamy ManikandanÎncă nu există evaluări
- Lachmapure2017Document6 paginiLachmapure2017Palanisamy ManikandanÎncă nu există evaluări
- 1 PDFDocument8 pagini1 PDFPalanisamy ManikandanÎncă nu există evaluări
- Monomeric C-Reactive Protein Aggravates Secondary Degeneration After Intracerebral Haemorrhagic Stroke and May Function As A Sensor For Systemic InflammationDocument12 paginiMonomeric C-Reactive Protein Aggravates Secondary Degeneration After Intracerebral Haemorrhagic Stroke and May Function As A Sensor For Systemic InflammationPalanisamy ManikandanÎncă nu există evaluări
- Exophiala Dermatitidis Endophthalmitis: Case Report and Literature ReviewDocument7 paginiExophiala Dermatitidis Endophthalmitis: Case Report and Literature ReviewPalanisamy ManikandanÎncă nu există evaluări
- Food Companies (November)Document15 paginiFood Companies (November)ITZMERZS100% (3)
- 14 PDFDocument21 pagini14 PDFPalanisamy ManikandanÎncă nu există evaluări
- Monomeric C-Reactive Protein Aggravates Secondary Degeneration After Intracerebral Haemorrhagic Stroke and May Function As A Sensor For Systemic InflammationDocument12 paginiMonomeric C-Reactive Protein Aggravates Secondary Degeneration After Intracerebral Haemorrhagic Stroke and May Function As A Sensor For Systemic InflammationPalanisamy ManikandanÎncă nu există evaluări
- Optimisation of The Method of Stem Cells Mediated Tissue Remodelling in Diabetic Wound HealingDocument4 paginiOptimisation of The Method of Stem Cells Mediated Tissue Remodelling in Diabetic Wound HealingPalanisamy ManikandanÎncă nu există evaluări
- In Vitro Susceptibility of Filamentous Fungi to Azole DrugsDocument6 paginiIn Vitro Susceptibility of Filamentous Fungi to Azole DrugsPalanisamy ManikandanÎncă nu există evaluări
- Find farm equipment in TamilnaduDocument5 paginiFind farm equipment in TamilnadudowÎncă nu există evaluări
- Jamson 2019Document9 paginiJamson 2019Palanisamy ManikandanÎncă nu există evaluări
- Guideline On Writing Learning Outcomes 2019Document6 paginiGuideline On Writing Learning Outcomes 2019Palanisamy ManikandanÎncă nu există evaluări
- Find farm equipment in TamilnaduDocument5 paginiFind farm equipment in TamilnadudowÎncă nu există evaluări
- Kredics Et Al 2015Document18 paginiKredics Et Al 2015Palanisamy ManikandanÎncă nu există evaluări
- Sreekumar Et Al PDFDocument7 paginiSreekumar Et Al PDFPalanisamy ManikandanÎncă nu există evaluări
- Essential for Health: Safe Drinking WaterDocument1 paginăEssential for Health: Safe Drinking WaterPalanisamy ManikandanÎncă nu există evaluări
- Polybacterial Community Analysis of Human Conjunctiva Reveals Novel BacteriaDocument12 paginiPolybacterial Community Analysis of Human Conjunctiva Reveals Novel BacteriaPalanisamy ManikandanÎncă nu există evaluări
- Merck PyrotoxinDocument6 paginiMerck PyrotoxinPalanisamy ManikandanÎncă nu există evaluări
- Essential for Health: Safe Drinking WaterDocument1 paginăEssential for Health: Safe Drinking WaterPalanisamy ManikandanÎncă nu există evaluări
- Manikandan Et AlDocument10 paginiManikandan Et AlPalanisamy ManikandanÎncă nu există evaluări
- Quality CentreDocument2 paginiQuality CentrePalanisamy ManikandanÎncă nu există evaluări
- Exophiala Dermatitidis Endophthalmitis: Case Report and Literature ReviewDocument7 paginiExophiala Dermatitidis Endophthalmitis: Case Report and Literature ReviewPalanisamy ManikandanÎncă nu există evaluări
- Manikandan Et AlDocument10 paginiManikandan Et AlPalanisamy ManikandanÎncă nu există evaluări
- ESCI 101 Discovering Emerging Global Literature 050818Document27 paginiESCI 101 Discovering Emerging Global Literature 050818Palanisamy ManikandanÎncă nu există evaluări
- FSSAI Regulation 2011Document7 paginiFSSAI Regulation 2011Palanisamy ManikandanÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Agarose Gel Electrophoresis (Full Report)Document10 paginiAgarose Gel Electrophoresis (Full Report)El LisÎncă nu există evaluări
- Lyphochek Assayed Chemistry Control Levels 1 and 2: Revision Date 2019-03-07 Indicates Revised InformationDocument1 paginăLyphochek Assayed Chemistry Control Levels 1 and 2: Revision Date 2019-03-07 Indicates Revised InformationReymart John GallardoÎncă nu există evaluări
- CRISPR Book Chapter PDFDocument21 paginiCRISPR Book Chapter PDFlepo sandra100% (1)
- Abbas Et Al. 2020 - Nature Immunology-2020-Abbas-TomaselloDocument33 paginiAbbas Et Al. 2020 - Nature Immunology-2020-Abbas-TomaselloAchille BroggiÎncă nu există evaluări
- Lab ReportDocument10 paginiLab ReportJeevikaGoyal100% (1)
- Introduction To Bioinformatics Lecture 3Document20 paginiIntroduction To Bioinformatics Lecture 3HaMmAd KhAnÎncă nu există evaluări
- Synthesis On Genetically Modified OrganismDocument3 paginiSynthesis On Genetically Modified OrganismSamsonÎncă nu există evaluări
- Stem Cell BiotechnologyDocument106 paginiStem Cell BiotechnologyTej deep saiÎncă nu există evaluări
- 1097-Article Text-1717-1-10-20190622Document9 pagini1097-Article Text-1717-1-10-20190622Nur HasanahÎncă nu există evaluări
- Crosstalk Among Plant Growth Regulators and Signaling MoleculesDocument3 paginiCrosstalk Among Plant Growth Regulators and Signaling MoleculesasifÎncă nu există evaluări
- Lab 4 DIFFERENTIAL & Special STAINDocument21 paginiLab 4 DIFFERENTIAL & Special STAINAAAÎncă nu există evaluări
- Molecular Weight MarkersDocument12 paginiMolecular Weight MarkersZecaValerianoÎncă nu există evaluări
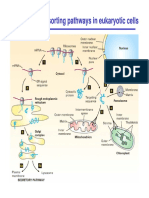
- Major Protein Sorting Pathways in Eukaryotic CellsDocument19 paginiMajor Protein Sorting Pathways in Eukaryotic CellsRara Aulia IIÎncă nu există evaluări
- BACTERIAL TOXINS: TARGETS AND MECHANISMSDocument20 paginiBACTERIAL TOXINS: TARGETS AND MECHANISMSYogiabrorÎncă nu există evaluări
- Test Bank Microbiology Evolving Science 3rd Edition Slonczewski FosterDocument12 paginiTest Bank Microbiology Evolving Science 3rd Edition Slonczewski FosterAmber Oneill100% (22)
- Laboratory Test Result Reference Value Description Unit Molecular and GenomicsDocument1 paginăLaboratory Test Result Reference Value Description Unit Molecular and GenomicsSszhÎncă nu există evaluări
- BiochemistryDocument7 paginiBiochemistryAbdelwahab AliÎncă nu există evaluări
- Genotoxicity of Drugs Mechanisms, Testing Guidelines and Methods For EvaluationDocument8 paginiGenotoxicity of Drugs Mechanisms, Testing Guidelines and Methods For Evaluationyogesh ushirÎncă nu există evaluări
- PHD Information Brochure 2021 22 Even 02-12-21Document13 paginiPHD Information Brochure 2021 22 Even 02-12-21Gmv VijuÎncă nu există evaluări
- Niacin and NAD+ DeficiencyDocument19 paginiNiacin and NAD+ DeficiencykatzrayÎncă nu există evaluări
- Hybridoma Technology: Production of Monoclonal AntibodiesDocument16 paginiHybridoma Technology: Production of Monoclonal AntibodiesDhanush BharadwajÎncă nu există evaluări
- PCR Primer Design GuidelinesDocument33 paginiPCR Primer Design Guidelinesendale gebregzabherÎncă nu există evaluări
- Chapter 28 Regulation of Gene Expression: Multiple Choice QuestionsDocument11 paginiChapter 28 Regulation of Gene Expression: Multiple Choice QuestionsKhristian Dom Ruam Cadabona40% (5)
- Fluoxetine Treatment For More Than 2 Days Decreases The Cellular Neuroplasticity and Microtubule Plasticity in Differentiated PC12 CellsDocument27 paginiFluoxetine Treatment For More Than 2 Days Decreases The Cellular Neuroplasticity and Microtubule Plasticity in Differentiated PC12 CellsRed DiggerÎncă nu există evaluări
- +2nd - Biochemical Engineering and Biotechnology - Prof. NajafpourDocument8 pagini+2nd - Biochemical Engineering and Biotechnology - Prof. NajafpourcikpayungÎncă nu există evaluări
- Emerging COVID-19 Variants and Their Impact On SARS-CoV-2 Diagnosis Therapeutics and VaccinesDocument18 paginiEmerging COVID-19 Variants and Their Impact On SARS-CoV-2 Diagnosis Therapeutics and VaccinesDoddy KurniawanÎncă nu există evaluări
- Assimilation of Digested FoodDocument8 paginiAssimilation of Digested FoodArash Halim83% (6)
- (PDF) Genetic Engineering Principles, Procedures and ConsequencesDocument27 pagini(PDF) Genetic Engineering Principles, Procedures and ConsequencesBhavani Patil0% (1)
- Lecture Notes - Weeks 1 and 2 BCMB2001Document17 paginiLecture Notes - Weeks 1 and 2 BCMB2001Anushka NeupaneÎncă nu există evaluări
- Chapter 8 Practice TestDocument6 paginiChapter 8 Practice Testcrqin qinÎncă nu există evaluări