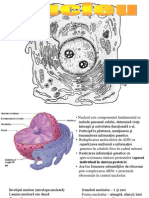
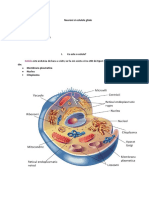
S-ar putea să vă placă și
- NUCLEUL1Document15 paginiNUCLEUL1PetrnelaÎncă nu există evaluări
- LP6 NucleuDocument15 paginiLP6 NucleuMihail RoscaÎncă nu există evaluări
- Tema 2. Nucleul PDFDocument6 paginiTema 2. Nucleul PDFAlina PinzaruÎncă nu există evaluări
- Nucleul CelularDocument7 paginiNucleul Celularcandi7Încă nu există evaluări
- Biologie Celulara LPDocument5 paginiBiologie Celulara LPRebeca FlorinaÎncă nu există evaluări
- Biologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiDocument31 paginiBiologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiMiruna-CristianaBirtuÎncă nu există evaluări
- NUCLEULDocument71 paginiNUCLEULAndrada Raicu100% (1)
- Nucleul Si Reproducerea Celulara Curs 2012 2013Document12 paginiNucleul Si Reproducerea Celulara Curs 2012 2013Bejenaru GeorgeÎncă nu există evaluări
- Document 5Document3 paginiDocument 5Good GuyÎncă nu există evaluări
- Nucleul CeluleiDocument34 paginiNucleul CeluleiElena PruniciÎncă nu există evaluări
- Nucleul CelularDocument25 paginiNucleul CelularGrațiela CazanÎncă nu există evaluări
- Nucleolul Şi Funcţiile LuiDocument2 paginiNucleolul Şi Funcţiile LuiNutza Zadic100% (1)
- Curs 4 BiocelDocument26 paginiCurs 4 BiocelBianca TomeciÎncă nu există evaluări
- CELULE Si CROMOZOMI Referat AnatomieDocument20 paginiCELULE Si CROMOZOMI Referat AnatomieLapadat MihaelaÎncă nu există evaluări
- 1.noţiuni Generale Despre CeluleDocument5 pagini1.noţiuni Generale Despre CeluleAndreea100% (1)
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- Nucleul - Cap. 6Document5 paginiNucleul - Cap. 6Nicolae IaşanÎncă nu există evaluări
- NucleulDocument3 paginiNucleuldaytayÎncă nu există evaluări
- Curs 9 Nucleul Si CromatinaDocument23 paginiCurs 9 Nucleul Si CromatinaTurturea ClaudiuÎncă nu există evaluări
- BioCel Curs 2 Nucleul Partea IDocument10 paginiBioCel Curs 2 Nucleul Partea IMarin TheodorÎncă nu există evaluări
- Curs 1 Bio MolecularaDocument123 paginiCurs 1 Bio MolecularaAmalia MoldovanuÎncă nu există evaluări
- Mio LogieDocument80 paginiMio LogieAmal AzkÎncă nu există evaluări
- BIOLOGIE CELULARA ReferatDocument19 paginiBIOLOGIE CELULARA Referatpraporgicu mirelaÎncă nu există evaluări
- Nucleul CelularDocument49 paginiNucleul CelularRaduBercaÎncă nu există evaluări
- 7 - NucleulDocument4 pagini7 - NucleulCiobanu MihaiÎncă nu există evaluări
- Celula EucariotaDocument15 paginiCelula Eucariotaiancu alexandru100% (2)
- Curs Genetică 2 UMFCD 2017-2018Document6 paginiCurs Genetică 2 UMFCD 2017-2018StefanChistolÎncă nu există evaluări
- NUCLEULDocument3 paginiNUCLEULMihai CodinÎncă nu există evaluări
- NUCLEULDocument43 paginiNUCLEULBondreaTiberiuÎncă nu există evaluări
- Curs Celula 2Document50 paginiCurs Celula 2Madalina Elena Răciulă cas RusuÎncă nu există evaluări
- Curs - Nucleul CelularDocument49 paginiCurs - Nucleul CelularOvidiu DumitruÎncă nu există evaluări
- Nucleul CursDocument11 paginiNucleul CursFekete VictoriaÎncă nu există evaluări
- NUCLEULDocument3 paginiNUCLEULcristi123321Încă nu există evaluări
- Replicarea ADNDocument11 paginiReplicarea ADNIulia100% (1)
- 73 e 5 Dfa 1Document4 pagini73 e 5 Dfa 1rovanaÎncă nu există evaluări
- Nucleul - ,,creierul CeluleiDocument33 paginiNucleul - ,,creierul CeluleiDana Semen100% (3)
- Nucleul - Notite de CursDocument19 paginiNucleul - Notite de CursMorgan Rosewood100% (1)
- Referat Anatomie-Nucleul CelularDocument7 paginiReferat Anatomie-Nucleul CelularAdina GheorgheÎncă nu există evaluări
- Proiect La Biologie: Clasaa9aD Paraschiv Seline AndreeaDocument7 paginiProiect La Biologie: Clasaa9aD Paraschiv Seline Andreea٭BUNNYÎncă nu există evaluări
- Neuroni Si Celule GlialeDocument8 paginiNeuroni Si Celule GlialeElenaÎncă nu există evaluări
- NUCLEULDocument9 paginiNUCLEULley_lyÎncă nu există evaluări
- Biologie CelularaDocument193 paginiBiologie CelularaAndrei GOÎncă nu există evaluări
- proiect bio clasa a 9-aDocument4 paginiproiect bio clasa a 9-abadeaionutdavid19Încă nu există evaluări
- CitogeneticaDocument0 paginiCitogeneticaGeorge BrinzeaÎncă nu există evaluări
- NeuronulDocument5 paginiNeuronulmiki.elaÎncă nu există evaluări
- Cap I - 2 - Elemente de Genetica MolecularaDocument45 paginiCap I - 2 - Elemente de Genetica MolecularaBogdan ButilcaÎncă nu există evaluări
- Organizarea Materialului GeneticDocument4 paginiOrganizarea Materialului GeneticmireaÎncă nu există evaluări
- Genetica MolecularaDocument3 paginiGenetica MolecularaIonuț BuzatuÎncă nu există evaluări
- Celula - Sinteza BACDocument9 paginiCelula - Sinteza BACAndreea Dinita100% (1)
- LP4 - Nucleul in InterfazaDocument4 paginiLP4 - Nucleul in InterfazaAna Rosu100% (1)
- Elemente de Biologie Celulară Şi MolecularăDocument196 paginiElemente de Biologie Celulară Şi MolecularăvalysaÎncă nu există evaluări
- Echipa RadiopozitivăDocument83 paginiEchipa RadiopozitivăTudor OstopoviciÎncă nu există evaluări
- Morfopat Imagini 20171853753727479404057Document153 paginiMorfopat Imagini 20171853753727479404057Nicoleta BodrugÎncă nu există evaluări
- Traumatismele Cranio CerebraleDocument28 paginiTraumatismele Cranio CerebraleTudor Ostopovici100% (1)
- Proiect-ILUMINAREA STRADALADocument6 paginiProiect-ILUMINAREA STRADALATudor OstopoviciÎncă nu există evaluări
- Scaunul in B InfDocument1 paginăScaunul in B InfTudor OstopoviciÎncă nu există evaluări
- Urograma 63976Document90 paginiUrograma 63976Tudor OstopoviciÎncă nu există evaluări
- Coxartrozareabilitare MedicalaDocument48 paginiCoxartrozareabilitare MedicalaTudor OstopoviciÎncă nu există evaluări
- Comparative Analysis of The Food Service Market in The Republic of Moldova and Other CountriesDocument8 paginiComparative Analysis of The Food Service Market in The Republic of Moldova and Other CountriesTudor OstopoviciÎncă nu există evaluări
- Intrebari Examen Dermato RezolvateDocument74 paginiIntrebari Examen Dermato RezolvateTudor OstopoviciÎncă nu există evaluări
- Sistemul Endocrin La VarstniciDocument9 paginiSistemul Endocrin La VarstnicinickklaussÎncă nu există evaluări
- Comparative Analysis of The Food Service Market in The Republic of Moldova and Other CountriesDocument8 paginiComparative Analysis of The Food Service Market in The Republic of Moldova and Other CountriesTudor OstopoviciÎncă nu există evaluări
- Proiect-ILUMINAREA STRADALADocument6 paginiProiect-ILUMINAREA STRADALATudor OstopoviciÎncă nu există evaluări
- Caz 8Document3 paginiCaz 8Кристи ПрипаÎncă nu există evaluări
- Fișă Pneumonie ComunitaraDocument8 paginiFișă Pneumonie ComunitaraTudor OstopoviciÎncă nu există evaluări
- Dezavantajele HomeopatieiDocument1 paginăDezavantajele HomeopatieiTudor OstopoviciÎncă nu există evaluări
- FJFJFHBJSJNDJBFJJNJNCJSJDocument25 paginiFJFJFHBJSJNDJBFJJNJNCJSJTudor OstopoviciÎncă nu există evaluări
- Scaunul in B InfDocument1 paginăScaunul in B InfTudor OstopoviciÎncă nu există evaluări
- Sinteze FisaDocument3 paginiSinteze FisaTudor OstopoviciÎncă nu există evaluări
- Caz ClinicDocument3 paginiCaz ClinicTudor OstopoviciÎncă nu există evaluări
- Caz Clinic SintezeDocument9 paginiCaz Clinic SintezeTudor OstopoviciÎncă nu există evaluări
- Cancerul ProfesionalDocument9 paginiCancerul ProfesionalCalin AndreeaÎncă nu există evaluări
- Sinteze FisaDocument3 paginiSinteze FisaTudor OstopoviciÎncă nu există evaluări
- Teste HelmintiDocument16 paginiTeste HelmintiTudor OstopoviciÎncă nu există evaluări
- 17 Teniaza - CisticercozaDocument2 pagini17 Teniaza - CisticercozaTudor OstopoviciÎncă nu există evaluări
- Anatomia Topografica A Mâinii PDFDocument58 paginiAnatomia Topografica A Mâinii PDFTudor OstopoviciÎncă nu există evaluări
- Regulile de Aplicare A Tratamentului Fitoterapeutic. Tratamentul Fitoterapeutic Al GastriteiDocument2 paginiRegulile de Aplicare A Tratamentului Fitoterapeutic. Tratamentul Fitoterapeutic Al GastriteiTudor OstopoviciÎncă nu există evaluări
- Cazuri CliniceDocument3 paginiCazuri CliniceTudor OstopoviciÎncă nu există evaluări
- LaserDocument2 paginiLaserTudor OstopoviciÎncă nu există evaluări
- Abcesul Si Flegmonul LaringeluiDocument1 paginăAbcesul Si Flegmonul LaringeluiCiorici StefanÎncă nu există evaluări
- Stenoza LaringianaDocument28 paginiStenoza LaringianaTudor OstopoviciÎncă nu există evaluări