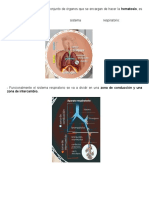
S-ar putea să vă placă și
- Solemnes de Bioquimica PDFDocument20 paginiSolemnes de Bioquimica PDFDavid CastilloÎncă nu există evaluări
- Taller Proteínas y AminoácidosDocument9 paginiTaller Proteínas y AminoácidosMarcoÎncă nu există evaluări
- Bq. Reacción de Transaminación y Su Reconocimiento Por Medio de Cromatografía en PapelDocument3 paginiBq. Reacción de Transaminación y Su Reconocimiento Por Medio de Cromatografía en PapelMarianne CortesÎncă nu există evaluări
- LipotropinaDocument58 paginiLipotropinaJosé Enrique Jurado Meneses50% (2)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De la EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Încă nu există evaluări
- Prueba Biología Transcripción y TraducciónDocument6 paginiPrueba Biología Transcripción y TraducciónLucas MoralesÎncă nu există evaluări
- Protocolo de CALIDADDocument39 paginiProtocolo de CALIDADMARIA JOSE ALEXANDRA AYALA OSORIOÎncă nu există evaluări
- Grupo 3 - Final de Biología UnlpDocument4 paginiGrupo 3 - Final de Biología UnlpjulianinaarandaokÎncă nu există evaluări
- Problemario Aa y ProteínasDocument8 paginiProblemario Aa y ProteínasalescrialferÎncă nu există evaluări
- Taller de EnzimasDocument5 paginiTaller de EnzimasKevin ParraÎncă nu există evaluări
- EXAMEN EXTRAORDINARIO - Martinez Segoviano Miguel AngelDocument5 paginiEXAMEN EXTRAORDINARIO - Martinez Segoviano Miguel AngelSangriento MartinezÎncă nu există evaluări
- Fosfatasa 1 y 2Document14 paginiFosfatasa 1 y 2carlosÎncă nu există evaluări
- Seminario Fisio GNRL 2023Document4 paginiSeminario Fisio GNRL 2023Valee FloresÎncă nu există evaluări
- Problemas BioquímicaDocument14 paginiProblemas BioquímicaElizabestÎncă nu există evaluări
- 02e Cinética EnzimáticaDocument95 pagini02e Cinética EnzimáticaFlor LopezÎncă nu există evaluări
- Examenes BioquimicaDocument71 paginiExamenes BioquimicaAranza Gutiérrez100% (1)
- Examen BCT 30 11 20 Prototipo BDocument12 paginiExamen BCT 30 11 20 Prototipo BNina SuarezÎncă nu există evaluări
- Prácticas 1-3 Fisiología GeneralDocument8 paginiPrácticas 1-3 Fisiología GeneralGaby RamosÎncă nu există evaluări
- Taller Sobre EnzimasDocument3 paginiTaller Sobre EnzimasDavid Julian Zambrano MorcilloÎncă nu există evaluări
- Preguntas ExamenDocument4 paginiPreguntas ExamenClara SerranoÎncă nu există evaluări
- Prototipo Primer Parcial BCT 09 05 2020Document6 paginiPrototipo Primer Parcial BCT 09 05 2020ESTIBALYZ JIMENEZ CASTA�EDAÎncă nu există evaluări
- Curva ProteinasDocument32 paginiCurva ProteinasBenjamin HonorioÎncă nu există evaluări
- Biomoléculas IDocument6 paginiBiomoléculas IJuanma AlfaroÎncă nu există evaluări
- Practica 5Document4 paginiPractica 5Tomas AsquiniÎncă nu există evaluări
- Diapositivas Enzimas IiDocument27 paginiDiapositivas Enzimas IiEVELYN ARIVILCA NINAÎncă nu există evaluări
- Resumen Fisio Prueba 1Document28 paginiResumen Fisio Prueba 1JavieraÎncă nu există evaluări
- Proteinas ImportanteDocument43 paginiProteinas ImportanteRuben MarquezÎncă nu există evaluări
- Taller EnzimasDocument4 paginiTaller EnzimasPaulaÎncă nu există evaluări
- Wuolah Free EXAMEN ORDINARIA BIOQUIMICADocument2 paginiWuolah Free EXAMEN ORDINARIA BIOQUIMICASara CanoreaÎncă nu există evaluări
- Wuolah Free EXAMEN ORDINARIA BIOQUIMICADocument2 paginiWuolah Free EXAMEN ORDINARIA BIOQUIMICASara CanoreaÎncă nu există evaluări
- IQI 313. Tercer Examen II 2020Document5 paginiIQI 313. Tercer Examen II 2020joel corsaÎncă nu există evaluări
- Clase Practica 2 Kenya Gil SDocument6 paginiClase Practica 2 Kenya Gil SKenya SantacruzÎncă nu există evaluări
- Practica Membrana CellDocument28 paginiPractica Membrana CellAlfredo BrenisÎncă nu există evaluări
- ProblemasDocument7 paginiProblemasJuce ChipanaÎncă nu există evaluări
- Prototipo Primer Parcial BCT PDFDocument6 paginiPrototipo Primer Parcial BCT PDFGege CuencaÎncă nu există evaluări
- Practicas 1 y 2 Quimica AnaliticaDocument6 paginiPracticas 1 y 2 Quimica AnaliticaJorge RamirezÎncă nu există evaluări
- Taller de Estudio. Proteína y EnzimasDocument7 paginiTaller de Estudio. Proteína y Enzimasmaria100% (1)
- Parcial 1Document6 paginiParcial 1Gabriel DannyÎncă nu există evaluări
- Problemario Bioquimica 2011Document15 paginiProblemario Bioquimica 2011Daniel PÎncă nu există evaluări
- LipotropinaDocument61 paginiLipotropinaJosé Enrique Jurado MenesesÎncă nu există evaluări
- Clase Tema 5a BiocatalisisDocument86 paginiClase Tema 5a BiocatalisisArianaRiosÎncă nu există evaluări
- Enzimas, Cofactores y VitaminasDocument14 paginiEnzimas, Cofactores y Vitaminasnatalia muñoz rodriguezÎncă nu există evaluări
- QUÍMICA ANALÍTICA I Intercambio IónicoDocument9 paginiQUÍMICA ANALÍTICA I Intercambio IónicoScott Thomas WeilandÎncă nu există evaluări
- Tema 9 FuncionCardiacaDocument12 paginiTema 9 FuncionCardiacaJoel Cristhian Rocabado GonzalesÎncă nu există evaluări
- Guía de Problemas 2 - Proteínas - LIGGA 2023Document3 paginiGuía de Problemas 2 - Proteínas - LIGGA 2023Gabriela LizamaÎncă nu există evaluări
- Tecnica MS MaditofDocument30 paginiTecnica MS MaditofCristina LizethÎncă nu există evaluări
- Autoevaluaciones EnzimasDocument29 paginiAutoevaluaciones Enzimasbelinchon96Încă nu există evaluări
- BC3123 Parcial I Prote Nas Yeisirhet Guerrero 17 10251Document6 paginiBC3123 Parcial I Prote Nas Yeisirhet Guerrero 17 10251Yeisireth Natalit Guerrero RodriguezÎncă nu există evaluări
- EeuvDocument6 paginiEeuvellechim2020Încă nu există evaluări
- El Objetivo de La Práctica Consiste en Aislar Una Proteína de Un Grupo de Ellas e IdentificarlaDocument8 paginiEl Objetivo de La Práctica Consiste en Aislar Una Proteína de Un Grupo de Ellas e IdentificarlalesperteyuÎncă nu există evaluări
- ALCANOSDocument18 paginiALCANOSJaime Aguilar50% (2)
- Preguntas de La 19 A La 22Document5 paginiPreguntas de La 19 A La 22jesus bravoÎncă nu există evaluări
- TP N 7 Operon Lac Giudici AbrilDocument8 paginiTP N 7 Operon Lac Giudici AbrilAbril GiudiciÎncă nu există evaluări
- DBIO 1076 2023-10 Guia Actividades 2 Unidad 1Document5 paginiDBIO 1076 2023-10 Guia Actividades 2 Unidad 1Juan Chávez SalazarÎncă nu există evaluări
- TGL Electroforesis ActDocument9 paginiTGL Electroforesis ActtoorreespvÎncă nu există evaluări
- Problemas AaDocument2 paginiProblemas AaflpdejuanÎncă nu există evaluări
- Enzimas e InhibiciónDocument73 paginiEnzimas e InhibiciónHéctor GalvánÎncă nu există evaluări
- BioM. Test 2.2Document6 paginiBioM. Test 2.2mq7tgdnk6rÎncă nu există evaluări
- Biosíntesis de ColesterolDocument6 paginiBiosíntesis de ColesterolAlejandra ViteriÎncă nu există evaluări
- Clase Practica 12 Kenya Gil SDocument5 paginiClase Practica 12 Kenya Gil SKenya SantacruzÎncă nu există evaluări
- Clase 1 - Biol272Document38 paginiClase 1 - Biol272Javier Pacheco BritoÎncă nu există evaluări
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De la EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Încă nu există evaluări
- Automatismo Cardíaco y Su RegulaciónDocument3 paginiAutomatismo Cardíaco y Su RegulaciónLucas MoralesÎncă nu există evaluări
- Fisio Respiratorio EurhesDocument17 paginiFisio Respiratorio EurhesLucas MoralesÎncă nu există evaluări
- Inervación Apuntes CoscarelliDocument4 paginiInervación Apuntes CoscarelliLucas MoralesÎncă nu există evaluări
- Comentario Película Strip SearchDocument1 paginăComentario Película Strip SearchLucas MoralesÎncă nu există evaluări
- Resumen Biología.Document5 paginiResumen Biología.Lucas MoralesÎncă nu există evaluări
- Informe 4 Laboratorio QuímicaDocument7 paginiInforme 4 Laboratorio Químicacesar HBÎncă nu există evaluări
- Materia Organica e InorganicaDocument3 paginiMateria Organica e InorganicaDelv NpÎncă nu există evaluări
- Propiedades de Los MaterialesDocument5 paginiPropiedades de Los Materialesgean villaltaÎncă nu există evaluări
- No. 8 MicroDocument8 paginiNo. 8 MicroMerryÎncă nu există evaluări
- QUIMICA PilasDocument15 paginiQUIMICA Pilasirebella18Încă nu există evaluări
- Pavimentos FlexiblesDocument2 paginiPavimentos FlexiblesJhobani Vasquez VelasquezÎncă nu există evaluări
- QUIZ 1 Corregido TermodinámicaDocument7 paginiQUIZ 1 Corregido TermodinámicaTracy KnowlesÎncă nu există evaluări
- Informe Solido-Liquido Extracción de CaféDocument15 paginiInforme Solido-Liquido Extracción de CaféJenni Lopez100% (1)
- CAPITULO 26 Tratamiento de LodosDocument15 paginiCAPITULO 26 Tratamiento de LodosFrancis LiberatoÎncă nu există evaluări
- Maquinaria para La Elaboracion de EmbutidosDocument6 paginiMaquinaria para La Elaboracion de EmbutidosYAnethÎncă nu există evaluări
- 3ro 3 Kimberly Barja Poma - Ciencia y Tecnologia INFORME de INDAGACIONDocument6 pagini3ro 3 Kimberly Barja Poma - Ciencia y Tecnologia INFORME de INDAGACIONLeticiaÎncă nu există evaluări
- 20 Cultivo de Hongos en La CasaDocument88 pagini20 Cultivo de Hongos en La CasapablobobaÎncă nu există evaluări
- Actualización de Ionomero de Vidrio 9Document23 paginiActualización de Ionomero de Vidrio 9Carmen Iturriaga Guajardo100% (1)
- DocumentologiaDocument8 paginiDocumentologialaloÎncă nu există evaluări
- 1.protocolo Administracion de MedicamentosDocument34 pagini1.protocolo Administracion de MedicamentosJUAN ESTEBANÎncă nu există evaluări
- Quispe-Rodriguez Examen Final - Trabajo Final de InvestigaciónDocument63 paginiQuispe-Rodriguez Examen Final - Trabajo Final de Investigaciónpatricia cieza torresÎncă nu există evaluări
- Termodinámica - Alejandro Vera Lázaro - 1edDocument30 paginiTermodinámica - Alejandro Vera Lázaro - 1edBryan Inca Enriquez75% (4)
- Libro 1Document1 paginăLibro 1LYDA ZAMIR LUENGASÎncă nu există evaluări
- Diapositivas PanDocument29 paginiDiapositivas PanErika Julissa0% (1)
- Tema+1.+Introduccion+ V1.1Document19 paginiTema+1.+Introduccion+ V1.1AndresD.MendozaÎncă nu există evaluări
- Grupo 232005 1 Tarea 2Document23 paginiGrupo 232005 1 Tarea 2Julian DavidÎncă nu există evaluări
- MSDS 000000000030488572 EsDocument9 paginiMSDS 000000000030488572 EsJennifer LondoñoÎncă nu există evaluări
- Qa - 04. - Cubiertas Planas Encuentros ParamentosDocument6 paginiQa - 04. - Cubiertas Planas Encuentros ParamentosRicardoVarelaÎncă nu există evaluări
- Tarea 3 Ejercicio 3 - Nicolas Alvarez Diagrama MentefactoDocument1 paginăTarea 3 Ejercicio 3 - Nicolas Alvarez Diagrama MentefactoDiasarmaÎncă nu există evaluări
- Informe PracticasDocument44 paginiInforme PracticasYonathan Aqepucho SapacayoÎncă nu există evaluări
- Analisis Instrumental en Calidad Del AireDocument30 paginiAnalisis Instrumental en Calidad Del AireYomar Garcia TrujilloÎncă nu există evaluări
- Libro MycomDocument97 paginiLibro MycomCesar Gabriel Carrasco MejiaÎncă nu există evaluări
- Tarea 5 SolidosDocument2 paginiTarea 5 SolidosMario Enrique Morales LemusÎncă nu există evaluări
- Metabolismo 2Document25 paginiMetabolismo 2Hernan DmgzÎncă nu există evaluări