S-ar putea să vă placă și
- P9 Elite 20221025Document4 paginiP9 Elite 20221025Chai AlvarezÎncă nu există evaluări
- Fetal Circulation ORIGINALDocument13 paginiFetal Circulation ORIGINALombir007Încă nu există evaluări
- Patterns of Metabolic Adaptation For Preterm and Term in 1st Neonatal WeekDocument10 paginiPatterns of Metabolic Adaptation For Preterm and Term in 1st Neonatal WeekEN Feb 22 UIÎncă nu există evaluări
- Higher Maternal Serum Prolactin Levels Are Associated With Reduced Glucose Tolerance During PregnancyDocument4 paginiHigher Maternal Serum Prolactin Levels Are Associated With Reduced Glucose Tolerance During Pregnancy06 Bhawandeep singhÎncă nu există evaluări
- Gastro 1Document15 paginiGastro 1Deddy WijayaÎncă nu există evaluări
- Raised PI Article AJOGDocument10 paginiRaised PI Article AJOGUmber AgarwalÎncă nu există evaluări
- Glucose profiles in healthy newbornsDocument12 paginiGlucose profiles in healthy newbornsEmilio Emmanué Escobar CruzÎncă nu există evaluări
- HHS Public Access: Gestational Diabetes MellitusDocument19 paginiHHS Public Access: Gestational Diabetes MellitusAnne MarieÎncă nu există evaluări
- A Prospective Study of The Relationships Between Movement and Glycemic Control During Day and Night in Pregnancyscientific ReportsDocument10 paginiA Prospective Study of The Relationships Between Movement and Glycemic Control During Day and Night in Pregnancyscientific ReportsMyrza AkbariÎncă nu există evaluări
- Role of Glycated Albumin During Pregnancy: Albumin Glikat Pada KehamilanDocument3 paginiRole of Glycated Albumin During Pregnancy: Albumin Glikat Pada KehamilanAditya PrabawaÎncă nu există evaluări
- 4396-20802-1-PB (Efek Insulin)Document5 pagini4396-20802-1-PB (Efek Insulin)FikriakbarÎncă nu există evaluări
- Plasma Levels and Placental Expression of Vaspin in Pregnant Women With Diabetes MellitusDocument7 paginiPlasma Levels and Placental Expression of Vaspin in Pregnant Women With Diabetes MellitusBala ChandiranÎncă nu există evaluări
- Urinary Congophilia in Women With Hypertensive Disorders of Pregnancy and Preexisting Proteinuria or HypertensionDocument7 paginiUrinary Congophilia in Women With Hypertensive Disorders of Pregnancy and Preexisting Proteinuria or HypertensionRyan AndrianÎncă nu există evaluări
- Poster Session II: ConclusionDocument1 paginăPoster Session II: ConclusionjuniorebindaÎncă nu există evaluări
- 2010 Update On Gestational DiabetesDocument13 pagini2010 Update On Gestational DiabetesAde Gustina SiahaanÎncă nu există evaluări
- Preeclampsia and Cognitive Impairment Later in Life - 2017 - American Journal of Obstetrics and Gynecology PDFDocument11 paginiPreeclampsia and Cognitive Impairment Later in Life - 2017 - American Journal of Obstetrics and Gynecology PDFfujimeisterÎncă nu există evaluări
- Hipoglicemia 4Document2 paginiHipoglicemia 4juanÎncă nu există evaluări
- Mir Ghani 2006Document4 paginiMir Ghani 2006Sarly FebrianaÎncă nu există evaluări
- Fructose Intake and Renal DiseaseDocument1 paginăFructose Intake and Renal DiseaseZebraÎncă nu există evaluări
- Paediatrica Indonesiana: Original ArticleDocument6 paginiPaediatrica Indonesiana: Original ArticleAyulita DianadewiÎncă nu există evaluări
- Diabetes Gestacional 2022 NejmDocument12 paginiDiabetes Gestacional 2022 NejmGabrielaÎncă nu există evaluări
- Sucrose-Rich Diet during Pregnancy Causes a Similar Response in Sprague-Dawley andDocument7 paginiSucrose-Rich Diet during Pregnancy Causes a Similar Response in Sprague-Dawley andmineortizvÎncă nu există evaluări
- Глюкагон мен глюкоза, инсулинDocument5 paginiГлюкагон мен глюкоза, инсулинArsen IzbanovÎncă nu există evaluări
- Probiotic supplements lower insulin resistance in GDMDocument8 paginiProbiotic supplements lower insulin resistance in GDMNana MuktiÎncă nu există evaluări
- Antenatal Corticosteroids and Perinatal Outcome in Late Fetal Growth Restriction: Analysis of Prospective CohortDocument7 paginiAntenatal Corticosteroids and Perinatal Outcome in Late Fetal Growth Restriction: Analysis of Prospective CohortYến ĐàoÎncă nu există evaluări
- Clinical Study of Infants of Diabetic MotherDocument112 paginiClinical Study of Infants of Diabetic Mothervighnaharta Online ServicesÎncă nu există evaluări
- Exploring The Relationship Between Preterm Placental Calcification and Adverse Maternal and Fetal OutcomeDocument7 paginiExploring The Relationship Between Preterm Placental Calcification and Adverse Maternal and Fetal OutcomeMaulida HalimahÎncă nu există evaluări
- Control de Glicemia en Embarazo PubmedDocument14 paginiControl de Glicemia en Embarazo PubmedCamila Pérez ArteagaÎncă nu există evaluări
- Be Hall 2006Document6 paginiBe Hall 2006adÎncă nu există evaluări
- Bowe, 2018)Document7 paginiBowe, 2018)Mayra PereiraÎncă nu există evaluări
- Ogtt 5Document4 paginiOgtt 5Arslan MubarikÎncă nu există evaluări
- Consensus criteria for diagnosing GDM following landmark HAPO studyDocument6 paginiConsensus criteria for diagnosing GDM following landmark HAPO studyRaul Garcia Salas CastilloÎncă nu există evaluări
- Investigación Cualitativa (Artículo 10)Document8 paginiInvestigación Cualitativa (Artículo 10)Wendy RamírezÎncă nu există evaluări
- Influence of Maternal Weight Gain On Birth Weight: A Gestational Diabetes CohortDocument9 paginiInfluence of Maternal Weight Gain On Birth Weight: A Gestational Diabetes Cohortمالك مناصرةÎncă nu există evaluări
- Placenta Changes in Pregnancy With Gestational Diabetes: R I G I N A L A P E RDocument6 paginiPlacenta Changes in Pregnancy With Gestational Diabetes: R I G I N A L A P E RMayra PereiraÎncă nu există evaluări
- Gestational Diabetes Induction of LaborDocument13 paginiGestational Diabetes Induction of LaborRalph AlbertoÎncă nu există evaluări
- Diabetes in Pregnancy JNDocument8 paginiDiabetes in Pregnancy JNpuspoÎncă nu există evaluări
- Domestic Animal EndocrinologyDocument9 paginiDomestic Animal EndocrinologyJose Luis Ponce CovarrubiasÎncă nu există evaluări
- Clinical and Molecular Characterisation of Hyperinsulinaemic Hypoglicemia in Infants Born Small For Gestation AgesDocument5 paginiClinical and Molecular Characterisation of Hyperinsulinaemic Hypoglicemia in Infants Born Small For Gestation AgesJaka KurniawanÎncă nu există evaluări
- Gupta 2020Document2 paginiGupta 2020dr dakshataÎncă nu există evaluări
- Practical Neonatal Endocrinology (PDFDrive)Document230 paginiPractical Neonatal Endocrinology (PDFDrive)ِAllya Al DabbaghÎncă nu există evaluări
- New Insights Into Gestational Glucose Metabolism: Lessons Learned From 21st Century ApproachesDocument8 paginiNew Insights Into Gestational Glucose Metabolism: Lessons Learned From 21st Century ApproachessanthiyasandyÎncă nu există evaluări
- 4 - Fetalneonatal 2014 307410Document7 pagini4 - Fetalneonatal 2014 307410Karina Mega WÎncă nu există evaluări
- Screening For Fetal Growth Restriction Using Ultrasound and The Sflt1/Plgf Ratio in Nulliparous Women: A Prospective Cohort StudyDocument13 paginiScreening For Fetal Growth Restriction Using Ultrasound and The Sflt1/Plgf Ratio in Nulliparous Women: A Prospective Cohort StudynicolasdlcaÎncă nu există evaluări
- Fetal Growth in Insulin Dependent DMDocument5 paginiFetal Growth in Insulin Dependent DMGeorge Tirta DihatmoÎncă nu există evaluări
- Oke MantapDocument7 paginiOke MantapMuhammad FadhilÎncă nu există evaluări
- A Lower Maternal Cortisol-to-Cortisone Ratio Precedes Clinical Diagnosis of Preterm and Term Preeclampsia by Many WeeksDocument12 paginiA Lower Maternal Cortisol-to-Cortisone Ratio Precedes Clinical Diagnosis of Preterm and Term Preeclampsia by Many WeeksMichael HusainÎncă nu există evaluări
- Nejmoa 2111673Document11 paginiNejmoa 2111673Vincent OsorioÎncă nu există evaluări
- Malaria in Pregnancy: The Difficulties in Measuring BirthweightDocument8 paginiMalaria in Pregnancy: The Difficulties in Measuring BirthweightEffika YuliaÎncă nu există evaluări
- Lactancia MaternaDocument6 paginiLactancia MaternaFernandaÎncă nu există evaluări
- Nutrition The Foetus and The Newly: NaismithDocument7 paginiNutrition The Foetus and The Newly: NaismithPierumberto PampaninÎncă nu există evaluări
- PQCNC 2019 Maternal/Newborn Initiative Kickoff - Neonatal Hypoglycemia and Outcomes: Are You Confused Yet?Document60 paginiPQCNC 2019 Maternal/Newborn Initiative Kickoff - Neonatal Hypoglycemia and Outcomes: Are You Confused Yet?kcochran100% (1)
- Visfatin and GDMDocument6 paginiVisfatin and GDMBala ChandiranÎncă nu există evaluări
- PhysioEx Exercise 9 Activity 5Document3 paginiPhysioEx Exercise 9 Activity 5Gonzeilo11Încă nu există evaluări
- Lipid Profile and Preeclampsia RiskDocument4 paginiLipid Profile and Preeclampsia Riskriki fazar rahmaditaÎncă nu există evaluări
- Jurnal PerinaDocument7 paginiJurnal PerinaBhismo PasetyoÎncă nu există evaluări
- Early Gestational Diabetes Screening in Obese Women: A Randomized Controlled TrialDocument8 paginiEarly Gestational Diabetes Screening in Obese Women: A Randomized Controlled Trialunie cantiqÎncă nu există evaluări
- Chapter 47 - HypoglycemiaDocument7 paginiChapter 47 - HypoglycemiaRafa HernandezÎncă nu există evaluări
- Maternal Insulin Treatment May Reduce Obesity Risk in OffspringDocument4 paginiMaternal Insulin Treatment May Reduce Obesity Risk in OffspringSarly FebrianaÎncă nu există evaluări
- Lindsay2015 PDFDocument11 paginiLindsay2015 PDFFelicia HalimÎncă nu există evaluări
- Correlations Between Parameters of Glycaemic Variability and Foetal Growth Neonatal Hypoglycaemia and Hyperbilirubinemia in Women With Gestational diabetesPLoS ONEDocument14 paginiCorrelations Between Parameters of Glycaemic Variability and Foetal Growth Neonatal Hypoglycaemia and Hyperbilirubinemia in Women With Gestational diabetesPLoS ONEVicente Fasce OlmosÎncă nu există evaluări
- Utilization of Nutrients During Postnatal Development: International Series of Monographs in Pure and Applied Biology: ZoologyDe la EverandUtilization of Nutrients During Postnatal Development: International Series of Monographs in Pure and Applied Biology: ZoologyÎncă nu există evaluări
- VFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZDocument1 paginăVFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZAbdullah QayyumÎncă nu există evaluări
- VFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZDocument1 paginăVFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZAbdullah QayyumÎncă nu există evaluări
- VFBDFNBFDBDBGN GGBGCDocument1 paginăVFBDFNBFDBDBGN GGBGCAbdullah QayyumÎncă nu există evaluări
- VFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZDocument1 paginăVFBDFNBFDBDBGNGFNBDGFBDFB DFBDF: F NCBVDDXDDVDFVSVVDVDVDSVDVZAbdullah QayyumÎncă nu există evaluări
- VFBDFNBFDBDBGN GGBGCDocument1 paginăVFBDFNBFDBDBGN GGBGCAbdullah QayyumÎncă nu există evaluări
- Secondary Headache: By: Mutia Dewi Assifa Supervisor: Dr. Riki Sukiandra, SP.SDocument60 paginiSecondary Headache: By: Mutia Dewi Assifa Supervisor: Dr. Riki Sukiandra, SP.SAbdullah QayyumÎncă nu există evaluări
- Case SarafDocument44 paginiCase SarafAbdullah QayyumÎncă nu există evaluări
- Headache Among Patient With HIV DeseaseDocument12 paginiHeadache Among Patient With HIV DeseaseAbdullah QayyumÎncă nu există evaluări
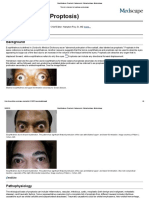
- Exophthalmos (Proptosis) - Background, Pathophysiology, EpidemiologyDocument7 paginiExophthalmos (Proptosis) - Background, Pathophysiology, EpidemiologyAbdullah QayyumÎncă nu există evaluări
- Headache Among Patient With HIV DeseaseDocument12 paginiHeadache Among Patient With HIV DeseaseAbdullah QayyumÎncă nu există evaluări
- Headache in Hiv Patient DiagnosticDocument3 paginiHeadache in Hiv Patient DiagnosticAbdullah QayyumÎncă nu există evaluări
- Human Complement Syntesis During Fetla LifeDocument9 paginiHuman Complement Syntesis During Fetla LifeAbdullah QayyumÎncă nu există evaluări
- Translate SarafDocument16 paginiTranslate SarafAbdullah QayyumÎncă nu există evaluări
- Congenital HipotirodismDocument17 paginiCongenital HipotirodismAbdullah QayyumÎncă nu există evaluări
- Daftar PustakaDocument5 paginiDaftar PustakaAbdullah QayyumÎncă nu există evaluări
- Fetal Growth and Development During PregnancyDocument13 paginiFetal Growth and Development During PregnancyJamica Anne Rias CostinianoÎncă nu există evaluări
- Infertility in Farm AnimalsDocument20 paginiInfertility in Farm AnimalsИгор ГалоскиÎncă nu există evaluări
- Giving Birth: Learning Objectives: After The Discussion of This Topic, You Should Be Able ToDocument126 paginiGiving Birth: Learning Objectives: After The Discussion of This Topic, You Should Be Able Tochesca_paunganÎncă nu există evaluări
- Carski Rez Po VejnovicuDocument48 paginiCarski Rez Po VejnovicuAndreyKovacevicÎncă nu există evaluări
- Chapt. 16 UtzDocument42 paginiChapt. 16 UtzMonicaÎncă nu există evaluări
- OBII - 16 Postterm Pregnancy - PDF Version 1Document6 paginiOBII - 16 Postterm Pregnancy - PDF Version 1Felina CabadingÎncă nu există evaluări
- Chapter 4 Maternal PhysiologyDocument12 paginiChapter 4 Maternal PhysiologyRem AlfelorÎncă nu există evaluări
- Annual Report 2009Document175 paginiAnnual Report 2009accord123Încă nu există evaluări
- Complications of Mitral Stenosis in PregnancyDocument3 paginiComplications of Mitral Stenosis in PregnancyJulissapenacÎncă nu există evaluări
- A Case of Suspected Illegal Abortion: How Clinicians May Assist The Forensic PathologistDocument43 paginiA Case of Suspected Illegal Abortion: How Clinicians May Assist The Forensic Pathologistjeremy ludwigÎncă nu există evaluări
- Factors Affecting Maternal Perception of Fetal MovementsDocument9 paginiFactors Affecting Maternal Perception of Fetal MovementsNoraÎncă nu există evaluări
- Psychological Perspective of Abnormal LaborDocument25 paginiPsychological Perspective of Abnormal LaborPriscilla Sarah PayneÎncă nu există evaluări
- Prenatal Development Stages & ProcessesDocument7 paginiPrenatal Development Stages & ProcessesRomel Ciptoadi WijayaÎncă nu există evaluări
- Hipoglicemia 2020 - Ingles PDFDocument18 paginiHipoglicemia 2020 - Ingles PDFMargaret MaggiÎncă nu există evaluări
- A Case of Brain Death During PregnancyDocument52 paginiA Case of Brain Death During PregnancyMa Pilar Peñaflor AñonuevoÎncă nu există evaluări
- The PlacentaDocument3 paginiThe PlacentaLiezelÎncă nu există evaluări
- Full Download Test Bank For Neonatal and Pediatric Respiratory Care 5th Edition by Walsh PDF Full ChapterDocument35 paginiFull Download Test Bank For Neonatal and Pediatric Respiratory Care 5th Edition by Walsh PDF Full Chapterlacrosseteosintejdwg100% (19)
- Test Bank For Maternity and Pediatric Nursing 2nd Edition Susan Scott RicciDocument7 paginiTest Bank For Maternity and Pediatric Nursing 2nd Edition Susan Scott RicciJulieJacobsrkozw100% (69)
- Pregnancy Metabolic Adaptation and Nutritional Requirements PDFDocument8 paginiPregnancy Metabolic Adaptation and Nutritional Requirements PDFMuh AfdholÎncă nu există evaluări
- Fetal CirculationDocument10 paginiFetal CirculationprembarnabasÎncă nu există evaluări
- Case-Study (3) JovenarDocument12 paginiCase-Study (3) JovenarJovenar MonsantoÎncă nu există evaluări
- Imminen Uterine Rupture: DR Arvid Tardan, SpogDocument18 paginiImminen Uterine Rupture: DR Arvid Tardan, SpogsiscaÎncă nu există evaluări
- Umbilical Cord ProlapseDocument17 paginiUmbilical Cord ProlapseAnggi Rezki Argha Nauli0% (1)
- Neonatal AscariasisDocument3 paginiNeonatal AscariasisAnonymous Tr4aGt7Încă nu există evaluări
- Infant of A Diabetic MotherDocument17 paginiInfant of A Diabetic MotherLovina Falendini AndriÎncă nu există evaluări
- Product Monograph Atripla (Emtricitabine - Tenofovir D. F. - Efavirenz)Document70 paginiProduct Monograph Atripla (Emtricitabine - Tenofovir D. F. - Efavirenz)tartaruga_tartarugaÎncă nu există evaluări
- Drugs Used in PregnancyDocument11 paginiDrugs Used in PregnancyTasmia RhamanÎncă nu există evaluări
- Midterm Exam Review for Maternal and Child NursingDocument7 paginiMidterm Exam Review for Maternal and Child NursingCres Padua QuinzonÎncă nu există evaluări