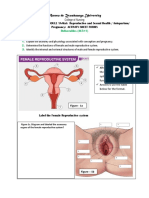
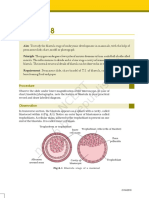
S-ar putea să vă placă și
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5795)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1091)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Paramount ClaimDocument2 paginiParamount ClaimCurry, William Lawrence III, agent100% (5)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- 9SELECT Mitosis vs. Meiosis Comparison Recap by Amoeba SistersDocument2 pagini9SELECT Mitosis vs. Meiosis Comparison Recap by Amoeba SistersArvie Jane BateÎncă nu există evaluări
- PCB3103C Exam 2 (Take-Home)Document5 paginiPCB3103C Exam 2 (Take-Home)Pmorris1898Încă nu există evaluări
- Perpetuaion of Life - How Animals ReproduceDocument3 paginiPerpetuaion of Life - How Animals ReproduceBenjamin Fernandez Jr.Încă nu există evaluări
- Superoksida Dismutase (Sod) Dan Radikal BebasDocument6 paginiSuperoksida Dismutase (Sod) Dan Radikal BebasIvon PalenteÎncă nu există evaluări
- 6.3 Meiosis Exam QuestionsDocument6 pagini6.3 Meiosis Exam QuestionsShae50% (2)
- Q.1 The Proximal End of The Filament of A Stamen Is Attached To The (A) Thalamus (B) Petal of The Flower (C) Anther (D) Both (A) and (B)Document95 paginiQ.1 The Proximal End of The Filament of A Stamen Is Attached To The (A) Thalamus (B) Petal of The Flower (C) Anther (D) Both (A) and (B)Priya SoniÎncă nu există evaluări
- Human Reproductive SystemDocument5 paginiHuman Reproductive SystemSubatomoÎncă nu există evaluări
- Asynchronous Activity Sheet FormsDocument8 paginiAsynchronous Activity Sheet FormsNur SanaaniÎncă nu există evaluări
- MitosisDocument4 paginiMitosis2003rahulcreatorÎncă nu există evaluări
- Integrated Principles of Zoology 16th Edition Hickman Test BankDocument21 paginiIntegrated Principles of Zoology 16th Edition Hickman Test Bankrobertykbxnd100% (22)
- Worksheet. 4. Asexual ReproductionDocument3 paginiWorksheet. 4. Asexual ReproductionLeonita SwandjajaÎncă nu există evaluări
- CAMP Response Element-Binding Protein (CREB) A Possible Signaling Molecule Link in The Pathophysiology of SchizophreniaDocument14 paginiCAMP Response Element-Binding Protein (CREB) A Possible Signaling Molecule Link in The Pathophysiology of SchizophreniaSofi MazzaropiÎncă nu există evaluări
- DLP - Biology F4 - Answers - Chap 15 (Final)Document5 paginiDLP - Biology F4 - Answers - Chap 15 (Final)JOSRIYN RATHNA A/P THAVARAJAN MoeÎncă nu există evaluări
- Week 6 Lecture 2 - Reproduction in Flowering Plant PDFDocument36 paginiWeek 6 Lecture 2 - Reproduction in Flowering Plant PDFAndy PikiÎncă nu există evaluări
- Science 8 Quarter 4 Module 2Document23 paginiScience 8 Quarter 4 Module 2eljentabzÎncă nu există evaluări
- Lelm 206Document5 paginiLelm 206Kritika MishraÎncă nu există evaluări
- Mammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsDocument12 paginiMammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsaufaaÎncă nu există evaluări
- Human Embryology: Presenting By: Ms.G.Nivedhapriya M, SC ., Nursing Sripms-Con CoimbatoreDocument74 paginiHuman Embryology: Presenting By: Ms.G.Nivedhapriya M, SC ., Nursing Sripms-Con CoimbatoreKavipriyaÎncă nu există evaluări
- INGLESDocument2 paginiINGLESdan VitheÎncă nu există evaluări
- 2.7 Transcription & TranslationDocument1 pagină2.7 Transcription & TranslationMELANNIE SOFIA ORTIZ CUBIDESÎncă nu există evaluări
- CH 7 Test Bank For Essential Cell Biology 3rd Edition AlbertsDocument20 paginiCH 7 Test Bank For Essential Cell Biology 3rd Edition AlbertsRokia GhariebÎncă nu există evaluări
- Epigeneration PDFDocument2 paginiEpigeneration PDFElgon Jaahiel BeyÎncă nu există evaluări
- Mitosis and Meiosis WebquestDocument3 paginiMitosis and Meiosis Webquestapi-3156932390% (1)
- Plant CycleDocument6 paginiPlant CycleMinistry of GamersÎncă nu există evaluări
- Gene Mutations-Essay 69Document2 paginiGene Mutations-Essay 69TharushiNethmiÎncă nu există evaluări
- Module 8 MeiosisDocument20 paginiModule 8 Meiosisapi-293001217100% (1)
- Petunjuk Kegiatan Praktikum IPADocument6 paginiPetunjuk Kegiatan Praktikum IPAAzzakia Oofy AurelieÎncă nu există evaluări
- One Gene One Enzyme TheoryDocument16 paginiOne Gene One Enzyme TheoryMuhammad ZakriaÎncă nu există evaluări
- Genetics PracticeDocument4 paginiGenetics PracticeYanaÎncă nu există evaluări