S-ar putea să vă placă și
- Taller Evaluativo 10 y 11Document2 paginiTaller Evaluativo 10 y 11Leidy PalechorÎncă nu există evaluări
- Ejercicios Sencillos Densidad - 3Document2 paginiEjercicios Sencillos Densidad - 3Leidy PalechorÎncă nu există evaluări
- Taller Evl 9Document1 paginăTaller Evl 9Leidy PalechorÎncă nu există evaluări
- Departamento de Física y QuímicaDocument1 paginăDepartamento de Física y QuímicaCristian Andres Vargas GuerreroÎncă nu există evaluări
- Nomenclatura StockDocument3 paginiNomenclatura StockLeidy PalechorÎncă nu există evaluări
- Ejercicios Sencillos DensidadDocument1 paginăEjercicios Sencillos DensidadYUZULY ANDREA AMAYA HERNANDEZÎncă nu există evaluări
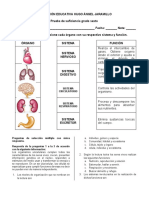
- Guia 1 - Sistema InmuneDocument5 paginiGuia 1 - Sistema InmuneLeidy PalechorÎncă nu există evaluări
- EVL - Origen Del Universo 6Document1 paginăEVL - Origen Del Universo 6Leidy PalechorÎncă nu există evaluări
- Evaluacion de SuficienciaDocument3 paginiEvaluacion de SuficienciaLeidy PalechorÎncă nu există evaluări
- Examen FINAL 9 QUIMICADocument1 paginăExamen FINAL 9 QUIMICALeidy PalechorÎncă nu există evaluări
- Tipos de Tejidos VegetalesDocument2 paginiTipos de Tejidos VegetalesLeidy PalechorÎncă nu există evaluări
- Evalyacion DensidadDocument2 paginiEvalyacion DensidadLeidy PalechorÎncă nu există evaluări
- Examen HomeostasisDocument1 paginăExamen HomeostasisLeidy PalechorÎncă nu există evaluări
- DetergentesDocument18 paginiDetergentesPedro SanchezÎncă nu există evaluări
- Labtrigliceridos19 1 PDFDocument3 paginiLabtrigliceridos19 1 PDFHéctor YaxÎncă nu există evaluări
- Haydee Dajhana Lòpez Ardila DB D1Document6 paginiHaydee Dajhana Lòpez Ardila DB D1Haydee LópezÎncă nu există evaluări
- Actualiza DoDocument11 paginiActualiza DoElizabeth AngelesÎncă nu există evaluări
- BiologiaDocument10 paginiBiologiaLEIDY MILAGROS ITO RAMOSÎncă nu există evaluări
- Control Resuelto Lípidos PDFDocument3 paginiControl Resuelto Lípidos PDFFiorella GiacomuzziÎncă nu există evaluări
- Quimica LecitinaDocument11 paginiQuimica LecitinaCecilia AlejandraÎncă nu există evaluări
- Formulación de Una EmulsiónDocument5 paginiFormulación de Una EmulsiónFabiola Galvan ValleÎncă nu există evaluări
- Elaboración de Productos de Aseo y CosmetologiaDocument10 paginiElaboración de Productos de Aseo y CosmetologiaDiomer Alzate BerrioÎncă nu există evaluări
- Resumen Ejecutivo de Un Proyecto de Innovación Tecnológica y Elección de Convocatoria de FinanciaciónDocument7 paginiResumen Ejecutivo de Un Proyecto de Innovación Tecnológica y Elección de Convocatoria de FinanciaciónMayra Olivo100% (1)
- Lipidos EnsayoDocument6 paginiLipidos EnsayoAnonymous fHB057lri100% (1)
- Metabolismo de Lipidos 2Document23 paginiMetabolismo de Lipidos 2cmtgpÎncă nu există evaluări
- Análisis FodaDocument89 paginiAnálisis FodaDogus DogusÎncă nu există evaluări
- Informe Glicerina Laboratorio de Quimica CuantitativaDocument17 paginiInforme Glicerina Laboratorio de Quimica CuantitativaGABRIEL AARON RIOS CARPIOÎncă nu există evaluări
- Producción de Propilenglicol (Oxido de Propileno)Document54 paginiProducción de Propilenglicol (Oxido de Propileno)Katherine Insuasty Martinez100% (1)
- Informe Perfil LipidicoDocument19 paginiInforme Perfil LipidicoLiliana Lino67% (3)
- Lípidos IIDocument110 paginiLípidos IIPatrick Corcino RetizÎncă nu există evaluări
- Anexo 1 - Carta Tecnológica Procesos de Transformación de Grasas y Aceites VegetalesDocument41 paginiAnexo 1 - Carta Tecnológica Procesos de Transformación de Grasas y Aceites VegetalesSofia MuñozÎncă nu există evaluări
- Tecnologías de Jabones, Grasas y Aceites - Wilmer Borras - Teoría Basica de JabonesDocument11 paginiTecnologías de Jabones, Grasas y Aceites - Wilmer Borras - Teoría Basica de JabonesEduardo ZuninoÎncă nu există evaluări
- Solventes No Acuosos en AguaDocument37 paginiSolventes No Acuosos en AguaLuciaÎncă nu există evaluări
- JABON LIQUIDO SabilaDocument20 paginiJABON LIQUIDO SabilaMetsy CabreraÎncă nu există evaluări
- Materia de BioquímicaDocument27 paginiMateria de BioquímicaFernando FiallosÎncă nu există evaluări
- Operaciones y Procesos UnitariosDocument121 paginiOperaciones y Procesos UnitariosPedro Chirinos MondragonÎncă nu există evaluări
- Cap 4Document5 paginiCap 4Laura CantúÎncă nu există evaluări
- Grupo de Alimentos Predominantemente Grasos - OdpDocument44 paginiGrupo de Alimentos Predominantemente Grasos - OdpBrahyan Leon IglesiasÎncă nu există evaluări
- Preparacion de Productos para El Hogar!Document37 paginiPreparacion de Productos para El Hogar!Luis Rodolfo Ramirez RojasÎncă nu există evaluări
- Complemento Lectivo Derivados Petroquimicos Del PropilenoDocument9 paginiComplemento Lectivo Derivados Petroquimicos Del PropilenoMarco Fernando Duich MedranoÎncă nu există evaluări
- FA GlicerolipidosDocument4 paginiFA GlicerolipidosPaulo Ernesto MarcheziÎncă nu există evaluări
- StripperDocument2 paginiStripperSofíaAlabartÎncă nu există evaluări
- Taller de Repaso 1.Document9 paginiTaller de Repaso 1.Freddy Andres Garcia AvilaÎncă nu există evaluări