S-ar putea să vă placă și
- Finite Difference, Finite Element, and Finite Volume Method: January 2005Document33 paginiFinite Difference, Finite Element, and Finite Volume Method: January 2005Amanuel GirmaÎncă nu există evaluări
- 2004 FDFEand FVmethodsfor PDEs Handbook Material ModellingDocument33 pagini2004 FDFEand FVmethodsfor PDEs Handbook Material ModellingAhmed KhalidÎncă nu există evaluări
- Comparison of Two Up-Scaling Methods in Poroelasticity and Its GeneralizationsDocument10 paginiComparison of Two Up-Scaling Methods in Poroelasticity and Its GeneralizationsAbhishek SharmaÎncă nu există evaluări
- Turing Patterns With Pentagonal SymmetryDocument9 paginiTuring Patterns With Pentagonal Symmetryricardoarmando.gonzalez2714Încă nu există evaluări
- American Association For The Advancement of ScienceDocument5 paginiAmerican Association For The Advancement of ScienceNamu PatelÎncă nu există evaluări
- Finite Difference, Finite Element, and Finite Volume Method: January 2005Document33 paginiFinite Difference, Finite Element, and Finite Volume Method: January 2005yohannesÎncă nu există evaluări
- Pattern Formation in A Generalized Chemotactic Model: Bulletin of Mathematical Biology (1998) 60Document26 paginiPattern Formation in A Generalized Chemotactic Model: Bulletin of Mathematical Biology (1998) 60Suman BhowmickÎncă nu există evaluări
- Reaction-Diffusion Model As A Framework For Understanding Biological Pattern FormationDocument6 paginiReaction-Diffusion Model As A Framework For Understanding Biological Pattern FormationbeemitsuÎncă nu există evaluări
- On Two Micromechanics Theories For Determining Micro Macro Relations in Heterogeneous SolidsDocument16 paginiOn Two Micromechanics Theories For Determining Micro Macro Relations in Heterogeneous Solidsdavibarbosa91Încă nu există evaluări
- Mat - 4Document23 paginiMat - 4Luis FuentesÎncă nu există evaluări
- Well-Posed and Ill-Posed Behaviour of The (I) - Rheology For Granular FlowDocument25 paginiWell-Posed and Ill-Posed Behaviour of The (I) - Rheology For Granular FlowRajkumar Chemical Engg. Dept.Încă nu există evaluări
- Asymptotic State Transformations of Continuous Variable ResourcesDocument61 paginiAsymptotic State Transformations of Continuous Variable Resourcesnawal.maftahÎncă nu există evaluări
- 2000 Physrevlet CrockerDocument4 pagini2000 Physrevlet CrockerMirza BasitÎncă nu există evaluări
- Perturbation Techniques and AsymptoticsDocument24 paginiPerturbation Techniques and AsymptoticsnadineÎncă nu există evaluări
- Turing's Theory of Morphogenesis: Where We Started, Where We Are and Where We Want To GoDocument17 paginiTuring's Theory of Morphogenesis: Where We Started, Where We Are and Where We Want To GoLEONARDO RAMÍREZÎncă nu există evaluări
- Connecting Theory To Experiment in Poroelasticity: Steven R. PrideDocument38 paginiConnecting Theory To Experiment in Poroelasticity: Steven R. PrideAbhishek SharmaÎncă nu există evaluări
- Assessing The Significance of The Correlation Between Two Spatial ProcessesDocument13 paginiAssessing The Significance of The Correlation Between Two Spatial ProcessesJoão Pedro CaetanoÎncă nu există evaluări
- Viscoelasticity RGDocument28 paginiViscoelasticity RGManu HegdeÎncă nu există evaluări
- Phase Equilibria in The Polydisperse Zwanzig Model of Hard RodsDocument14 paginiPhase Equilibria in The Polydisperse Zwanzig Model of Hard RodsGilberto RuizÎncă nu există evaluări
- Tensor AlgebraDocument8 paginiTensor AlgebraMaggyBalcazarÎncă nu există evaluări
- Numerical Calculation of Effective Density and Compressibility Tensors in Periodic Porous Media: A Multi-Scale Asymptotic MethodDocument6 paginiNumerical Calculation of Effective Density and Compressibility Tensors in Periodic Porous Media: A Multi-Scale Asymptotic MethodAkinbode Sunday OluwagbengaÎncă nu există evaluări
- 1 s2.0 S0378437112004025 MainDocument10 pagini1 s2.0 S0378437112004025 Mainargelio arroyoÎncă nu există evaluări
- Numerical Study of Segregation Using A New Drag Force Correlation For Polydisperse Systems Derived From Lattice-Boltzmann SimulationsDocument10 paginiNumerical Study of Segregation Using A New Drag Force Correlation For Polydisperse Systems Derived From Lattice-Boltzmann SimulationsArpit DwivediÎncă nu există evaluări
- An Optimization Algorithm Applied To The Morr 2007 International Journal ofDocument10 paginiAn Optimization Algorithm Applied To The Morr 2007 International Journal of姚志豪Încă nu există evaluări
- PilliodPuckett VOFDocument38 paginiPilliodPuckett VOFSomdeb BandopadhyayÎncă nu există evaluări
- An Interesting Class of Partial Differential Equations: 131.155.81.2 On: Sat, 22 Nov 2014 19:01:55Document22 paginiAn Interesting Class of Partial Differential Equations: 131.155.81.2 On: Sat, 22 Nov 2014 19:01:55Ricemilk JogjaÎncă nu există evaluări
- 5 - Hanotin 2015 - Viscoelasticity of Vibrated Granular SuspensionsDocument22 pagini5 - Hanotin 2015 - Viscoelasticity of Vibrated Granular SuspensionskarolgoodnotesÎncă nu există evaluări
- Generalized Lorenz Models Routes Chaos Energy ConservingDocument15 paginiGeneralized Lorenz Models Routes Chaos Energy ConservingValeria Montserrat Maya GonzalezÎncă nu există evaluări
- A Discrete Model For The Apparent Viscosity of Polydisperse Suspensions Including Maximum Packing FractionDocument14 paginiA Discrete Model For The Apparent Viscosity of Polydisperse Suspensions Including Maximum Packing FractionLuke ParryÎncă nu există evaluări
- PrintDocument23 paginiPrintMarcD2009Încă nu există evaluări
- Quantum Biology at The Cellular Level - Elements of The Research ProgramDocument49 paginiQuantum Biology at The Cellular Level - Elements of The Research ProgramGustavo EngenheiroÎncă nu există evaluări
- Marco Baldi Et Al - Hydrodynamical N-Body Simulations of Coupled Dark Energy CosmologiesDocument21 paginiMarco Baldi Et Al - Hydrodynamical N-Body Simulations of Coupled Dark Energy CosmologiesMuths999999Încă nu există evaluări
- The Equilibrium Behavior of Reversible Coagulation-Fragmentation ProcessesDocument28 paginiThe Equilibrium Behavior of Reversible Coagulation-Fragmentation ProcessesJanCarlBrionesÎncă nu există evaluări
- Cross-Diffusion Systems With Entropy Structure JungelDocument79 paginiCross-Diffusion Systems With Entropy Structure JungeljcgomezuÎncă nu există evaluări
- On Dense Granular Flows: T E P J EDocument25 paginiOn Dense Granular Flows: T E P J EOpu DebnathÎncă nu există evaluări
- Diffusion Model and Analysis of Diffusion Process at Lagrangian MethodDocument30 paginiDiffusion Model and Analysis of Diffusion Process at Lagrangian MethodZohaib AliÎncă nu există evaluări
- Maximum Likelihood Estimation of Limited and Discrete Dependent Variable Models With Nested Random EffectsDocument23 paginiMaximum Likelihood Estimation of Limited and Discrete Dependent Variable Models With Nested Random EffectsJaime NavarroÎncă nu există evaluări
- 2022 Singh NCRDDocument40 pagini2022 Singh NCRDSatyvir SinghÎncă nu există evaluări
- Mignone 2007 ApJS 170 228Document15 paginiMignone 2007 ApJS 170 228Nikhil Arun KumarÎncă nu există evaluări
- A Non-Standard Statistical Approach To The Silo Discharge: T E P JDocument8 paginiA Non-Standard Statistical Approach To The Silo Discharge: T E P JA94leo14hsetmaÎncă nu există evaluări
- Mittal1 PDFDocument28 paginiMittal1 PDFVedant DubeyÎncă nu există evaluări
- Diffusion4 PDFDocument17 paginiDiffusion4 PDFlolaÎncă nu există evaluări
- Stability Analysis and Consistent Solitary Wave Solutions For - 2023 - Results IDocument16 paginiStability Analysis and Consistent Solitary Wave Solutions For - 2023 - Results Ironaldquezada038Încă nu există evaluări
- 1997a+a 320 1BDocument7 pagini1997a+a 320 1Bga.simplicio2Încă nu există evaluări
- The Route To Self-Similarity in Turbulent Jets and Plumes: Byguillaumecarazzo, Edouardkaminski StephentaitDocument12 paginiThe Route To Self-Similarity in Turbulent Jets and Plumes: Byguillaumecarazzo, Edouardkaminski StephentaitZulfiqar AliÎncă nu există evaluări
- 2014 11 25 Mycek Et AlDocument30 pagini2014 11 25 Mycek Et AlFauzan S EÎncă nu există evaluări
- Parameter-Robust Preconditioners For Biot's ModelDocument30 paginiParameter-Robust Preconditioners For Biot's ModelAxel DorianÎncă nu există evaluări
- FDM, Fem, FVMDocument32 paginiFDM, Fem, FVMRaj EaswarmoorthiÎncă nu există evaluări
- 19 Aop1414Document38 pagini19 Aop1414singhmukul7410Încă nu există evaluări
- COMSOL Multiphysics For Efficient Solution of A TRDocument8 paginiCOMSOL Multiphysics For Efficient Solution of A TRMohamed Ould MoussaÎncă nu există evaluări
- Venturi JFMDocument23 paginiVenturi JFMgovindÎncă nu există evaluări
- Journal of The Mechanics and Physics of Solids: Noureddine Damil, Michel Potier-FerryDocument15 paginiJournal of The Mechanics and Physics of Solids: Noureddine Damil, Michel Potier-FerryBoumediene BakhaddaÎncă nu există evaluări
- Null Control KDV Coupled Bilinear SystemDocument31 paginiNull Control KDV Coupled Bilinear SystemJefferson Prada MárquezÎncă nu există evaluări
- Atmosphere: Concentration Fluctuations and Odor Dispersion in Lagrangian ModelsDocument23 paginiAtmosphere: Concentration Fluctuations and Odor Dispersion in Lagrangian ModelsDinku Seyoum ZelekeÎncă nu există evaluări
- Model SimulationDocument17 paginiModel SimulationMario Enrique Bocchio DiazÎncă nu există evaluări
- Quantum Biology at Cellular Level PDFDocument49 paginiQuantum Biology at Cellular Level PDFKalpit SahooÎncă nu există evaluări
- Entropy: Quantum and Ecosystem EntropiesDocument13 paginiEntropy: Quantum and Ecosystem EntropiesjohnnirbanoÎncă nu există evaluări
- Baranyi Et Al 1993 A Non-Autonomous Differential Equation To Model Bacterial Growth PDFDocument17 paginiBaranyi Et Al 1993 A Non-Autonomous Differential Equation To Model Bacterial Growth PDFCarlos AndradeÎncă nu există evaluări
- Granular Materials at Meso-scale: Towards a Change of Scale ApproachDe la EverandGranular Materials at Meso-scale: Towards a Change of Scale ApproachÎncă nu există evaluări
- Large-Order Behaviour of Perturbation TheoryDe la EverandLarge-Order Behaviour of Perturbation TheoryÎncă nu există evaluări
- p214-231 - Devolution of Health CentersDocument18 paginip214-231 - Devolution of Health CentersRuby GarciaÎncă nu există evaluări
- Due Diligence Tool For Savings and Credit CoopsDocument17 paginiDue Diligence Tool For Savings and Credit CoopsRuby GarciaÎncă nu există evaluări
- An Overview of Age-Friendly Cities and Communities Around The WorldDocument20 paginiAn Overview of Age-Friendly Cities and Communities Around The WorldRuby GarciaÎncă nu există evaluări
- Social Protection in The PhilippinesDocument12 paginiSocial Protection in The PhilippinesRuby GarciaÎncă nu există evaluări
- Pidspn9809 - Regulation of Deposit-Taking CooperativesDocument4 paginiPidspn9809 - Regulation of Deposit-Taking CooperativesRuby GarciaÎncă nu există evaluări
- Credit Union EmpowermentDocument14 paginiCredit Union EmpowermentRuby GarciaÎncă nu există evaluări
- Philippines Legal Framework Analysis National Report - CoopDocument26 paginiPhilippines Legal Framework Analysis National Report - CoopRuby GarciaÎncă nu există evaluări
- Resource Mobilization Action Plan UpdateDocument13 paginiResource Mobilization Action Plan UpdateRuby GarciaÎncă nu există evaluări
- Lao Challenges Social ProtectionDocument3 paginiLao Challenges Social ProtectionRuby GarciaÎncă nu există evaluări
- Resource Mobilization For CSOsDocument6 paginiResource Mobilization For CSOsRuby GarciaÎncă nu există evaluări
- Mobile Health Care ServiceDocument2 paginiMobile Health Care ServiceRuby GarciaÎncă nu există evaluări
- Theme Paper III Partnership and Resourcee MobilizationDocument10 paginiTheme Paper III Partnership and Resourcee MobilizationRuby GarciaÎncă nu există evaluări
- RRHO Training - Resource MobilizationDocument69 paginiRRHO Training - Resource MobilizationRuby GarciaÎncă nu există evaluări
- Strengthening The Referral Mechanism in A Service Delivery NetworkDocument12 paginiStrengthening The Referral Mechanism in A Service Delivery NetworkRuby Garcia100% (1)
- ASEAN at 30 - CambodiaDocument21 paginiASEAN at 30 - CambodiaRuby GarciaÎncă nu există evaluări
- OM Shanti - Moving Together in PeaceDocument15 paginiOM Shanti - Moving Together in PeaceRuby GarciaÎncă nu există evaluări
- Research Manual-ProceduresDocument18 paginiResearch Manual-ProceduresRuby GarciaÎncă nu există evaluări
- Poverty Issue Paper AgeingDocument5 paginiPoverty Issue Paper AgeingRuby GarciaÎncă nu există evaluări
- 15th-DGF-Proceedings - As-Of-30-Nov - 15 Years of DecentralizationDocument28 pagini15th-DGF-Proceedings - As-Of-30-Nov - 15 Years of DecentralizationRuby GarciaÎncă nu există evaluări
- The Catalyst Fund: A New Era in Evaluating The Reproductive Justice MovementDocument37 paginiThe Catalyst Fund: A New Era in Evaluating The Reproductive Justice MovementRuby GarciaÎncă nu există evaluări
- 01 - IRA ImprovementDocument124 pagini01 - IRA ImprovementRuby GarciaÎncă nu există evaluări
- SBP Notes-1 PDFDocument7 paginiSBP Notes-1 PDFzeeshanÎncă nu există evaluări
- Complete Cocker Spaniel Guide 009 PDFDocument119 paginiComplete Cocker Spaniel Guide 009 PDFElmo RÎncă nu există evaluări
- EEN 203 Slide Notes Year 2018: PART I - Numbers and CodesDocument78 paginiEEN 203 Slide Notes Year 2018: PART I - Numbers and CodesSHIVAM CHOPRAÎncă nu există evaluări
- Pre Joining Guidelines For InfosysDocument21 paginiPre Joining Guidelines For InfosysGokul0% (2)
- God's Word in Holy Citadel New Jerusalem" Monastery, Glodeni - Romania, Redactor Note. Translated by I.ADocument6 paginiGod's Word in Holy Citadel New Jerusalem" Monastery, Glodeni - Romania, Redactor Note. Translated by I.Abillydean_enÎncă nu există evaluări
- How To Use The ActionDocument3 paginiHow To Use The Actioncizgiaz cizgiÎncă nu există evaluări
- D. Michael Quinn-Same-Sex Dynamics Among Nineteenth-Century Americans - A MORMON EXAMPLE-University of Illinois Press (2001)Document500 paginiD. Michael Quinn-Same-Sex Dynamics Among Nineteenth-Century Americans - A MORMON EXAMPLE-University of Illinois Press (2001)xavirreta100% (3)
- The Trial of Jesus ChristDocument10 paginiThe Trial of Jesus ChristTomaso Vialardi di Sandigliano100% (2)
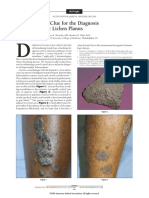
- A Novel Visual Clue For The Diagnosis of Hypertrophic Lichen PlanusDocument1 paginăA Novel Visual Clue For The Diagnosis of Hypertrophic Lichen Planus600WPMPOÎncă nu există evaluări
- BUS 301 - Hospitality Industry Vietnam - Nguyễn Thị Thanh Thuý - 1632300205Document55 paginiBUS 301 - Hospitality Industry Vietnam - Nguyễn Thị Thanh Thuý - 1632300205Nguyễn Thị Thanh ThúyÎncă nu există evaluări
- IN THE BEGINNING WAS AFFECT RolnikDocument22 paginiIN THE BEGINNING WAS AFFECT RolnikFabiana PaulinoÎncă nu există evaluări
- Review of Related LiteratureDocument9 paginiReview of Related LiteratureMarion Joy GanayoÎncă nu există evaluări
- Boylestad Circan 3ce Ch02Document18 paginiBoylestad Circan 3ce Ch02sherry mughalÎncă nu există evaluări
- Black Body RadiationDocument46 paginiBlack Body RadiationKryptosÎncă nu există evaluări
- Case 3 GROUP-6Document3 paginiCase 3 GROUP-6Inieco RacheleÎncă nu există evaluări
- Vocabulary Words - 20.11Document2 paginiVocabulary Words - 20.11ravindra kumar AhirwarÎncă nu există evaluări
- 7 CAAT-AIR-GM03 Guidance-Material-for-Foreign-Approved-Maintenance-Organization - I3R0 - 30oct2019 PDFDocument59 pagini7 CAAT-AIR-GM03 Guidance-Material-for-Foreign-Approved-Maintenance-Organization - I3R0 - 30oct2019 PDFJindarat KasemsooksakulÎncă nu există evaluări
- Especificação - PneusDocument10 paginiEspecificação - Pneusmarcos eduÎncă nu există evaluări
- ZultaniteDocument4 paginiZultaniteAcharya BalwantÎncă nu există evaluări
- Design and Experimental Performance Assessment of An Outer Rotor PM Assisted SynRM For The Electric Bike PropulsionDocument11 paginiDesign and Experimental Performance Assessment of An Outer Rotor PM Assisted SynRM For The Electric Bike PropulsionTejas PanchalÎncă nu există evaluări
- Signal&Systems - Lab Manual - 2021-1Document121 paginiSignal&Systems - Lab Manual - 2021-1telecom_numl8233Încă nu există evaluări
- Venue:: Alberta Electrical System Alberta Electrical System OperatorDocument48 paginiVenue:: Alberta Electrical System Alberta Electrical System OperatorOmar fethiÎncă nu există evaluări
- Tutorial Class 4: Finders As Bailee Right of A Bailee General LienDocument26 paginiTutorial Class 4: Finders As Bailee Right of A Bailee General Lienchirag jainÎncă nu există evaluări
- Green Biocomposites For Structural ApplicationsDocument27 paginiGreen Biocomposites For Structural ApplicationsLamia Nour Ben abdelrahmenÎncă nu există evaluări
- Komatsu Hydraulic Excavator Pc290lc 290nlc 6k Shop ManualDocument20 paginiKomatsu Hydraulic Excavator Pc290lc 290nlc 6k Shop Manualmallory100% (47)
- Title: Daily Visit Report: SDL Mini - Project Academic Year 2020-21 Group ID:GB6Document2 paginiTitle: Daily Visit Report: SDL Mini - Project Academic Year 2020-21 Group ID:GB6Arjuna JppÎncă nu există evaluări
- Surefire Hellfighter Power Cord QuestionDocument3 paginiSurefire Hellfighter Power Cord QuestionPedro VianaÎncă nu există evaluări
- COSL Brochure 2023Document18 paginiCOSL Brochure 2023DaniloÎncă nu există evaluări
- Bike Chasis DesignDocument7 paginiBike Chasis Designparth sarthyÎncă nu există evaluări
- Knowledge, Attitude and Practice of Non-Allied Health Sciences Students of Southwestern University Phinma During The Covid-19 PandemicDocument81 paginiKnowledge, Attitude and Practice of Non-Allied Health Sciences Students of Southwestern University Phinma During The Covid-19 Pandemicgeorgemayhew1030Încă nu există evaluări