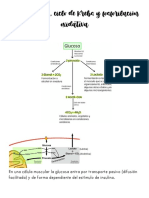
S-ar putea să vă placă și
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De la EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Încă nu există evaluări
- Respiracion CelularDocument8 paginiRespiracion CelularJeniffer ChaÎncă nu există evaluări
- Ciclo de Los Acidos Tricarboxilicos-2020Document52 paginiCiclo de Los Acidos Tricarboxilicos-2020SusanaÎncă nu există evaluări
- 12 Ciclo de Krebs Summary Lehninger Principles of Biochemistry PDFDocument14 pagini12 Ciclo de Krebs Summary Lehninger Principles of Biochemistry PDFJuli SaviniÎncă nu există evaluări
- ADA 2. Equipo 4. Piruvato DeshidrogenasaDocument28 paginiADA 2. Equipo 4. Piruvato DeshidrogenasaIvan Pech CauichÎncă nu există evaluări
- 16-CICLO DE ÁCIDO CITRICO ResumenDocument17 pagini16-CICLO DE ÁCIDO CITRICO ResumenCecilia Cea HenriquezÎncă nu există evaluări
- T2 Ciclo Del Ácido Cítrico - Ana2020-AlumnosDocument18 paginiT2 Ciclo Del Ácido Cítrico - Ana2020-AlumnosGustavo EimbckeÎncă nu există evaluări
- Descarboxilación Oxidativa Del Piruvato - Ciclo de KrebsDocument7 paginiDescarboxilación Oxidativa Del Piruvato - Ciclo de KrebsMariaÎncă nu există evaluări
- TEMA 5 Respiracion Celular y Fermentacion.Document5 paginiTEMA 5 Respiracion Celular y Fermentacion.mariaÎncă nu există evaluări
- El Complejo de La Piruvato Deshidrogenasa Necesita 5 CoenzimasDocument3 paginiEl Complejo de La Piruvato Deshidrogenasa Necesita 5 CoenzimasalezÎncă nu există evaluări
- Piruvato DeshidrogenasaDocument7 paginiPiruvato DeshidrogenasaM PÎncă nu există evaluări
- Informe MitocondriaDocument12 paginiInforme MitocondriaEvelyn NeiraÎncă nu există evaluări
- Complejo PDHDocument9 paginiComplejo PDHJanys GilÎncă nu există evaluări
- Ejercicios de Mitocondria y Respiración Celular.Document5 paginiEjercicios de Mitocondria y Respiración Celular.Diana Karen Butanda PuenteÎncă nu există evaluări
- Preguntas Metabolismo CelularDocument5 paginiPreguntas Metabolismo CelularJhessica JancoÎncă nu există evaluări
- Fosforilación Oxidativa y Cadena Transporte ElectronesDocument29 paginiFosforilación Oxidativa y Cadena Transporte ElectronesniamineÎncă nu există evaluări
- Cadena de Transporte de ElectronesDocument29 paginiCadena de Transporte de ElectronesMarina GrenierÎncă nu există evaluări
- MetabolismoDocument7 paginiMetabolismoFatima Serrano PerezÎncă nu există evaluări
- Bioquímica Ambiental Tema: Metabolismo Y Glucolisis Ing - Quím. Carlos AltamiranoDocument46 paginiBioquímica Ambiental Tema: Metabolismo Y Glucolisis Ing - Quím. Carlos AltamiranoLina RojoÎncă nu există evaluări
- Tema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Document15 paginiTema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Alexander SanchezÎncă nu există evaluări
- Tema 10. CatabolismoDocument16 paginiTema 10. CatabolismoAny RuizÎncă nu există evaluări
- Seminario 15 - Texto - Cadena de Transporte de Electrones. Fosforilación Oxidativa - 2021Document23 paginiSeminario 15 - Texto - Cadena de Transporte de Electrones. Fosforilación Oxidativa - 2021Joseph ReyesÎncă nu există evaluări
- Resumen 4Document47 paginiResumen 4Ramiro MartinÎncă nu există evaluări
- Bioquímica - T14Document37 paginiBioquímica - T14Maria Gomez SabaterÎncă nu există evaluări
- Complejo PDHDocument8 paginiComplejo PDHkarlaÎncă nu există evaluări
- Respiración Celular - GlosarioDocument3 paginiRespiración Celular - GlosarioCarlos ChiaÎncă nu există evaluări
- Cadena RespiratoriaDocument60 paginiCadena RespiratoriaYoselin RomeroÎncă nu există evaluări
- Tema 13Document11 paginiTema 13daliasandulacheÎncă nu există evaluări
- Seminario 13 - Cadena de Transporte de Electrones. Fosforilación Oxidativa - 2019Document24 paginiSeminario 13 - Cadena de Transporte de Electrones. Fosforilación Oxidativa - 2019Nati Di NucciÎncă nu există evaluări
- Fosforilación OxidativaDocument20 paginiFosforilación OxidativaValeri Vasquez Cucho0% (1)
- Bioquimica II, Bioenergetica y MetabolismoDocument40 paginiBioquimica II, Bioenergetica y MetabolismoOneida MorenoÎncă nu există evaluări
- MitocondriaDocument5 paginiMitocondriaKevin Bravo SanchezÎncă nu există evaluări
- UNIDAD 2 Temas 13 y 14Document38 paginiUNIDAD 2 Temas 13 y 14LuisÎncă nu există evaluări
- BiolooDocument11 paginiBiolooDani LoaizaÎncă nu există evaluări
- Etapas Del Ciclo de KrebsDocument13 paginiEtapas Del Ciclo de KrebsADry BenithezÎncă nu există evaluări
- Cuestionario Respiracion y FotosintesisDocument5 paginiCuestionario Respiracion y FotosintesisAnonymous 5RyfXG7100% (1)
- Tema 4Document6 paginiTema 4Maria GarciaÎncă nu există evaluări
- Ev 3. Presentación de Las Vías y Procesos Metabólicos Identificados en El Metabolismo AeróbicoDocument60 paginiEv 3. Presentación de Las Vías y Procesos Metabólicos Identificados en El Metabolismo AeróbicoAhovitÎncă nu există evaluări
- Ciclo de Krebs, Cadena Respiratoria y Fosforilación OxidativaDocument8 paginiCiclo de Krebs, Cadena Respiratoria y Fosforilación OxidativaJohan Fernando HerreraÎncă nu există evaluări
- Procesos Bioenergeticos A Nivel Mitocondrial - Semana 8 - KarolineDocument17 paginiProcesos Bioenergeticos A Nivel Mitocondrial - Semana 8 - KarolineKAROLINE FRANKSHESCA CRUZ TUMEÎncă nu există evaluări
- Ciclo de KrebsDocument6 paginiCiclo de KrebsHector MironÎncă nu există evaluări
- Ciclo de KrebsDocument9 paginiCiclo de KrebsYeanni CalorÎncă nu există evaluări
- Resolución Del Taller 1 - Introducción Al MetabolismoDocument11 paginiResolución Del Taller 1 - Introducción Al Metabolismossmrt.2016Încă nu există evaluări
- Tema 10 BioquímicaDocument3 paginiTema 10 BioquímicaSara Menéndez RoblesÎncă nu există evaluări
- Ciclo de KrebsDocument55 paginiCiclo de Krebschristian_giambellucÎncă nu există evaluări
- Metabolismo Gluco Kbres MedicinaDocument40 paginiMetabolismo Gluco Kbres MedicinaLuis eduardo La torre AdrianzénÎncă nu există evaluări
- Relación de La Glucosis y La Fosforilación OxidativaDocument4 paginiRelación de La Glucosis y La Fosforilación OxidativaEymi GarcíaÎncă nu există evaluări
- Resumen - Reacción Del Piruvato A Acetilcoa, Reacciones Del Ciclo de Los Ácidos TricarboxílicosDocument5 paginiResumen - Reacción Del Piruvato A Acetilcoa, Reacciones Del Ciclo de Los Ácidos Tricarboxílicosanmenc04Încă nu există evaluări
- Cadena RespiratoriaDocument4 paginiCadena RespiratoriaJoiss Chirinos100% (1)
- Respiración AerobiaDocument3 paginiRespiración AerobiaLuis ReyÎncă nu există evaluări
- METABOLISMODocument4 paginiMETABOLISMOJimena OcanaÎncă nu există evaluări
- Semana 7Document47 paginiSemana 7Marilyn TantaquispeÎncă nu există evaluări
- Bioquímica Tema 2 (Fase Común Metabolismo)Document27 paginiBioquímica Tema 2 (Fase Común Metabolismo)Karlos CaballeroÎncă nu există evaluări
- Ciclo de KrebsDocument80 paginiCiclo de KrebsJORGE EDUARDO PARKER100% (1)
- Respiracion Celular Unidad 10Document7 paginiRespiracion Celular Unidad 10valentinaÎncă nu există evaluări
- Cadena Respiratoria PDFDocument8 paginiCadena Respiratoria PDFSusana Calderón Toledo0% (1)
- Fisiologia Aplicada S05s1Document25 paginiFisiologia Aplicada S05s1Tatiana IngaÎncă nu există evaluări
- Las MitocondriasDocument7 paginiLas MitocondriasOsvaldo UrreloÎncă nu există evaluări
- 5 Conversiondelpiruvatoaco ADocument15 pagini5 Conversiondelpiruvatoaco Amichel cautiÎncă nu există evaluări
- Agua en Los Alimentos PDFDocument15 paginiAgua en Los Alimentos PDFFlorencia BarsiÎncă nu există evaluări
- Aceites y Grasas PDFDocument19 paginiAceites y Grasas PDFFlorencia BarsiÎncă nu există evaluări
- Fosforilacion OxidativaDocument16 paginiFosforilacion OxidativaFlorencia BarsiÎncă nu există evaluări
- Elementos de La Función RenalDocument8 paginiElementos de La Función RenalFlorencia BarsiÎncă nu există evaluări
- Aminoacidos, Peptidos y ProteinasDocument21 paginiAminoacidos, Peptidos y ProteinasFlorencia Barsi100% (2)
- Estructura Tridimensional de Las ProteinasDocument18 paginiEstructura Tridimensional de Las ProteinasFlorencia BarsiÎncă nu există evaluări
- Toxicologia AlimentariaDocument7 paginiToxicologia AlimentariaFlorencia Barsi0% (1)
- Sistema Nervioso SensorialDocument35 paginiSistema Nervioso SensorialFiorella Coro AlvaÎncă nu există evaluări
- Monografia TuberculosisDocument21 paginiMonografia TuberculosisMeliza Zuu Barboza Huanca100% (1)
- Inmunocromatografia de VIHDocument8 paginiInmunocromatografia de VIHEdgarÎncă nu există evaluări
- AntiácidoDocument21 paginiAntiácidoJulie DZ33% (3)
- Banco de Preguntas Dr. Villacis (Completo)Document46 paginiBanco de Preguntas Dr. Villacis (Completo)leonardoÎncă nu există evaluări
- Caso Clinico PCR ListoooooooooDocument81 paginiCaso Clinico PCR ListoooooooooAlicia Pamela Avila Beltran0% (1)
- FICHAC 1 FloresDocument27 paginiFICHAC 1 FloresSoporte SistemasÎncă nu există evaluări
- Examen OrtodonciaDocument3 paginiExamen OrtodonciacruzesmeraldaÎncă nu există evaluări
- Album de FarmacosDocument5 paginiAlbum de FarmacosCarlos50% (2)
- Actividad1 RiesgoDocument4 paginiActividad1 RiesgoSolmar Del Valle Peña RiveroÎncă nu există evaluări
- Estres Electroestimulacion Isr1Document48 paginiEstres Electroestimulacion Isr1bronzonÎncă nu există evaluări
- Ubicación de Los Electrodos Del EKGDocument10 paginiUbicación de Los Electrodos Del EKGJose HoyerÎncă nu există evaluări
- Fisiología Del Estomago 6-03-19Document14 paginiFisiología Del Estomago 6-03-19Carol Jennifer Núñez Campos100% (1)
- Hoja Procesos Engranaje CilindricoDocument3 paginiHoja Procesos Engranaje CilindricoPepe ChupinÎncă nu există evaluări
- Potencia Anaeróbica y Capacidad AeróbicaDocument13 paginiPotencia Anaeróbica y Capacidad AeróbicavictoriaÎncă nu există evaluări
- Abdomen Normal y PatológicoDocument135 paginiAbdomen Normal y PatológicoMara Karyme SuasteÎncă nu există evaluări
- Vias Piramidales Resumen EstudianteDocument5 paginiVias Piramidales Resumen EstudianteJaime GarciaÎncă nu există evaluări
- FORMATODocument9 paginiFORMATOVICTOR HUGO CEDILLO CHALACOÎncă nu există evaluări
- Ciencias Naturales 6°Document6 paginiCiencias Naturales 6°ingreethpeñaÎncă nu există evaluări
- Afasia EstimulacionDocument10 paginiAfasia EstimulacionertyujÎncă nu există evaluări
- Materia Prima Del PanetonDocument6 paginiMateria Prima Del PanetonRIDERÎncă nu există evaluări
- Anatomia de Pelvis y Miembro Inf 2022Document314 paginiAnatomia de Pelvis y Miembro Inf 2022GABRIELA DE LOS ANGELES GONZALEZ RUIZÎncă nu există evaluări
- Cuadro Comparativo Entre Hongos Virus y BacteriasDocument1 paginăCuadro Comparativo Entre Hongos Virus y Bacteriasjpps100% (6)
- Para Diagnosticar en OdontologiaDocument5 paginiPara Diagnosticar en OdontologiaRosa RiveraÎncă nu există evaluări
- Andreas Ludwig Kalcker - ParásitosDocument13 paginiAndreas Ludwig Kalcker - ParásitosAnonymous MXBabgkV100% (1)
- BiodescodificacionDocument46 paginiBiodescodificacionalegaba75100% (2)
- Rubrica para Evaluar Portafolio de Evidencias 2016 n300Document6 paginiRubrica para Evaluar Portafolio de Evidencias 2016 n300ALEJANDRO OSTOIC ROZZIÎncă nu există evaluări
- Practica #06 NematodosDocument36 paginiPractica #06 NematodosEd CotÎncă nu există evaluări
- Farmacia Natural 2Document16 paginiFarmacia Natural 2Kevin Paul PincayÎncă nu există evaluări
- Introduccion Al CorteDocument11 paginiIntroduccion Al CorteLuis GonzálezÎncă nu există evaluări