S-ar putea să vă placă și
- Fisio II (Guia 1)Document12 paginiFisio II (Guia 1)manuel herradoraÎncă nu există evaluări
- Tejido MuscularDocument5 paginiTejido MuscularRichard Rene Rivera RamirezÎncă nu există evaluări
- 4Document45 pagini4José DalaÎncă nu există evaluări
- Funcion de Los Musculos SuperficialesDocument56 paginiFuncion de Los Musculos SuperficialesMaria Acosta100% (3)
- Documento Completo - pdf-PDFA PDFDocument18 paginiDocumento Completo - pdf-PDFA PDFoscarbio2009Încă nu există evaluări
- Tejido MuscularDocument8 paginiTejido MuscularQuipyÎncă nu există evaluări
- Manual Avanzado Capacitación en Elaboración de CárnicosDocument149 paginiManual Avanzado Capacitación en Elaboración de CárnicosRolando GandolfoÎncă nu există evaluări
- Tejido MuscularDocument12 paginiTejido MuscularMaria Virginia Alvarado LopezÎncă nu există evaluări
- Miología GeneralDocument13 paginiMiología GeneralBrand A Bernal100% (1)
- Tejido MuscularDocument5 paginiTejido MuscularCris FernandezÎncă nu există evaluări
- Tarea: Histología Muscular: Sistema OsteomioarticularDocument5 paginiTarea: Histología Muscular: Sistema OsteomioarticularHaraviel RiveraÎncă nu există evaluări
- Tejido MúscularDocument5 paginiTejido Múscular7prw2vvcwvÎncă nu există evaluări
- Tejido MuscularDocument11 paginiTejido MuscularGraciela Romero PalacioÎncă nu există evaluări
- Cuestionario MúsculoDocument8 paginiCuestionario MúsculoDARWIN RUDY CAMPERO GUZMÁNÎncă nu există evaluări
- Taller Biologia Tejido Muscular y NerviosoDocument14 paginiTaller Biologia Tejido Muscular y NerviosopablochillefrasesÎncă nu există evaluări
- Constitucion Anatomica Del MusculoDocument5 paginiConstitucion Anatomica Del Musculotokito212Încă nu există evaluări
- AVO 7 Tejido MuscularJY LCXDocument16 paginiAVO 7 Tejido MuscularJY LCXAbril CarinoÎncă nu există evaluări
- Tema 10. Fisiología Del MúsculoDocument16 paginiTema 10. Fisiología Del MúsculoKarla GarritsenÎncă nu există evaluări
- Tejido Muscular Según La Evolución de Los AnimalesDocument12 paginiTejido Muscular Según La Evolución de Los AnimalesInes Yulisa oviedo zuñigaÎncă nu există evaluări
- Mus CuloDocument32 paginiMus Culomarib89Încă nu există evaluări
- Musculo Primera ParteDocument21 paginiMusculo Primera ParteCristian OrlegoÎncă nu există evaluări
- Sistema MuscularDocument9 paginiSistema MuscularCarlos GaetanÎncă nu există evaluări
- Seminario 3 MusculoDocument14 paginiSeminario 3 Musculomartha sofiaÎncă nu există evaluări
- Tejido MuscularDocument3 paginiTejido MuscularEsteban Ramos MarinÎncă nu există evaluări
- Tema 19 (Tejido Muscular)Document37 paginiTema 19 (Tejido Muscular)Carlos Daniel Cárdenas SantosÎncă nu există evaluări
- Tejido MuscularDocument4 paginiTejido Muscularmaria tiguaÎncă nu există evaluări
- Apuntes Tejido MuscularDocument18 paginiApuntes Tejido MuscularPaz GarciaÎncă nu există evaluări
- Tejido Muscular Material Complementario (Histología)Document12 paginiTejido Muscular Material Complementario (Histología)miguel bravoÎncă nu există evaluări
- Biofisica de La Contraccion MuscularDocument12 paginiBiofisica de La Contraccion MuscularRuthy Sugey García SánchezÎncă nu există evaluări
- Apuntes Tejido MuscularDocument11 paginiApuntes Tejido Muscularedugalindo8Încă nu există evaluări
- MANUAL DE LAB 3 Estudiantes 2023Document11 paginiMANUAL DE LAB 3 Estudiantes 2023Carlos Mauricio VizcarraÎncă nu există evaluări
- Histologia Del Tejido Muscular PDFDocument3 paginiHistologia Del Tejido Muscular PDFroberto carlodÎncă nu există evaluări
- Sistema MuscularDocument5 paginiSistema MuscularPauloMazziniÎncă nu există evaluări
- Tejido Muscular 21Document44 paginiTejido Muscular 21Duarte MarceÎncă nu există evaluări
- Brenda Tejido Muscular Por Dentro Del Titan ColosalDocument6 paginiBrenda Tejido Muscular Por Dentro Del Titan ColosalJesus Abel Quiñonez ValenzuelaÎncă nu există evaluări
- TEJIDO MUSCULAR Dra MaldonadoDocument8 paginiTEJIDO MUSCULAR Dra MaldonadoLoismer LinaresÎncă nu există evaluări
- Tejido Muscular Histologia 2019Document4 paginiTejido Muscular Histologia 2019Ricardo SalasÎncă nu există evaluări
- Músculo LisoDocument3 paginiMúsculo LisoLianGaming HDÎncă nu există evaluări
- Tejido MuscularDocument28 paginiTejido MuscularCHRISTIAN ALBERTO AVILA TORRESÎncă nu există evaluări
- Cuestionario de Tejido MuscularDocument5 paginiCuestionario de Tejido MuscularlinuxhouseÎncă nu există evaluări
- Tejido MuscularDocument31 paginiTejido MuscularLastJd100% (2)
- RESUMEN FISIOLOGIA 1 ExamenDocument101 paginiRESUMEN FISIOLOGIA 1 ExamenFavio Alexis DíazÎncă nu există evaluări
- Estructra y Funciones Del Muscculo EsqueleticoDocument13 paginiEstructra y Funciones Del Muscculo EsqueleticoMaricruz Cordova DuarteÎncă nu există evaluări
- Músculo y NervioDocument30 paginiMúsculo y NervioYamila CoceresÎncă nu există evaluări
- Tejido MuscularDocument5 paginiTejido MuscularJuliana ArdilaÎncă nu există evaluări
- Cuáles Son Los Tipos de Tejido MuscularDocument12 paginiCuáles Son Los Tipos de Tejido Muscularelfisico666Încă nu există evaluări
- Musculo 1Document24 paginiMusculo 1Rafael PazÎncă nu există evaluări
- Coordinación MotoraDocument10 paginiCoordinación MotoraKhrriz CastilloÎncă nu există evaluări
- Tejido MuscularDocument13 paginiTejido MuscularVale IsbarboÎncă nu există evaluări
- Cuestionario 6Document6 paginiCuestionario 6Brayan Francisco Quispe AtaÎncă nu există evaluări
- Lectura Fisiología de La Contracción MuscularDocument6 paginiLectura Fisiología de La Contracción MuscularSANTIAGO ANDRES PLAZAS MARTINEZÎncă nu există evaluări
- Clase 5. Sistema MuscularDocument17 paginiClase 5. Sistema Muscularluichigo 222Încă nu există evaluări
- Trabajo Histología MúsculoDocument12 paginiTrabajo Histología MúsculoDomeniqueÎncă nu există evaluări
- Sistema MuscularDocument24 paginiSistema Muscularkmacho4280Încă nu există evaluări
- Sistema Muscular de AnimalessDocument19 paginiSistema Muscular de AnimalessNelson CallesÎncă nu există evaluări
- Informe de Laboratorio 4. Grupo 6Document15 paginiInforme de Laboratorio 4. Grupo 6christian vera0% (1)
- Tejido MuscularDocument9 paginiTejido MuscularOscar PerezÎncă nu există evaluări
- Anatomía artística 5: Articulaciones y funciones muscularesDe la EverandAnatomía artística 5: Articulaciones y funciones muscularesEvaluare: 4 din 5 stele4/5 (1)
- Tableros de DistribuciónDocument2 paginiTableros de Distribuciónkatty niño llacsahuangaÎncă nu există evaluări
- Manual Losas de CimentacionDocument6 paginiManual Losas de Cimentacionkatty niño llacsahuangaÎncă nu există evaluări
- Pa Procedimientos Constructivos Aula A - Marco BrummertDocument9 paginiPa Procedimientos Constructivos Aula A - Marco Brummertkatty niño llacsahuangaÎncă nu există evaluări
- Manual Losas de CimentacionDocument6 paginiManual Losas de Cimentacionkatty niño llacsahuangaÎncă nu există evaluări
- Sesion 10 Lectura de Planos 15 Set 2021Document28 paginiSesion 10 Lectura de Planos 15 Set 2021katty niño llacsahuangaÎncă nu există evaluări
- Sesion 08 Cajas de CentroDocument27 paginiSesion 08 Cajas de Centrokatty niño llacsahuangaÎncă nu există evaluări
- TAREA N 03 - Bendezu - Ducastaing - NiñoDocument7 paginiTAREA N 03 - Bendezu - Ducastaing - Niñokatty niño llacsahuangaÎncă nu există evaluări
- Control de Calidad de Los AgregadoDocument1 paginăControl de Calidad de Los Agregadokatty niño llacsahuangaÎncă nu există evaluări
- Los VirusDocument5 paginiLos Viruskatty niño llacsahuangaÎncă nu există evaluări
- Introducción LuminotecniaDocument12 paginiIntroducción Luminotecniakatty niño llacsahuangaÎncă nu există evaluări
- Gestion de Calidad AgregadosDocument30 paginiGestion de Calidad Agregadoskatty niño llacsahuangaÎncă nu există evaluări
- PARTE 1, 4 y 5Document7 paginiPARTE 1, 4 y 5katty niño llacsahuangaÎncă nu există evaluări
- Introducción LuminotecniaDocument12 paginiIntroducción Luminotecniakatty niño llacsahuangaÎncă nu există evaluări
- PARTE 1, 4 y 5Document7 paginiPARTE 1, 4 y 5katty niño llacsahuangaÎncă nu există evaluări
- Pa Aocc - Fundamentos de Obras de Infraestructuras - Iib Noche - Marco BrummertDocument8 paginiPa Aocc - Fundamentos de Obras de Infraestructuras - Iib Noche - Marco Brummertkatty niño llacsahuangaÎncă nu există evaluări
- Exámenes Topo IDocument6 paginiExámenes Topo Ikatty niño llacsahuangaÎncă nu există evaluări
- Maqueta de La Plaza de Armas de La Provincia de AyabacaDocument9 paginiMaqueta de La Plaza de Armas de La Provincia de Ayabacakatty niño llacsahuangaÎncă nu există evaluări
- Laboratorio 1 MSIDocument7 paginiLaboratorio 1 MSIkatty niño llacsahuangaÎncă nu există evaluări
- MonografiaDocument11 paginiMonografiakatty niño llacsahuangaÎncă nu există evaluări
- Resumen de Los Siete Ensayos de Interpretacion de La Realidad PeruanaDocument6 paginiResumen de Los Siete Ensayos de Interpretacion de La Realidad PeruanaTiffany BatesÎncă nu există evaluări
- Laboratorio 1 MSIDocument7 paginiLaboratorio 1 MSIkatty niño llacsahuangaÎncă nu există evaluări
- Estadística N°02Document11 paginiEstadística N°02katty niño llacsahuangaÎncă nu există evaluări
- EstadisticaDocument6 paginiEstadisticakatty niño llacsahuangaÎncă nu există evaluări
- Práctica de CampoDocument3 paginiPráctica de Campokatty niño llacsahuangaÎncă nu există evaluări
- Práctica de CampoDocument3 paginiPráctica de Campokatty niño llacsahuangaÎncă nu există evaluări
- Diseño CampoDocument1 paginăDiseño Campokatty niño llacsahuangaÎncă nu există evaluări
- Inteligencia WordDocument21 paginiInteligencia Wordkatty niño llacsahuangaÎncă nu există evaluări
- Silabo GeologiaDocument3 paginiSilabo Geologiakatty niño llacsahuangaÎncă nu există evaluări
- Diapositivas de La Teoria de KantDocument16 paginiDiapositivas de La Teoria de Kantkatty niño llacsahuangaÎncă nu există evaluări
- Laboratorio de TM Primera ParteDocument7 paginiLaboratorio de TM Primera Partekatty niño llacsahuangaÎncă nu există evaluări
- Actualizacion Guia LepraDocument152 paginiActualizacion Guia LepraTatan Rodriguez BucheliÎncă nu există evaluări
- Aves de Europa - Lynx PDFDocument113 paginiAves de Europa - Lynx PDFDani Cruzado ZarzaÎncă nu există evaluări
- Proyecto Áulico de PracticaDocument4 paginiProyecto Áulico de PracticaVanesa BarberisÎncă nu există evaluări
- Recetas OrdenesDocument3 paginiRecetas OrdenesDario PaillachoÎncă nu există evaluări
- El León y El RatónDocument2 paginiEl León y El RatónDeivi Jacks Castro Benites80% (5)
- Caperucita Roja - Teresa Wilms MonttDocument3 paginiCaperucita Roja - Teresa Wilms MonttTania MunizagaÎncă nu există evaluări
- 3 Fisiología Del EritrocitoDocument36 pagini3 Fisiología Del EritrocitoGustavo JimenezÎncă nu există evaluări
- Tema 3 Conductas Reprod. Maria Penado 2017Document87 paginiTema 3 Conductas Reprod. Maria Penado 2017Raquel de la HeraÎncă nu există evaluări
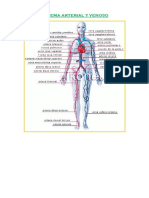
- Sistema Arterial y VenosoDocument12 paginiSistema Arterial y Venosoesthefanie ñontolÎncă nu există evaluări
- Tobacco and Alcohol Use in G-Rated Childrens Anim - En.esDocument7 paginiTobacco and Alcohol Use in G-Rated Childrens Anim - En.esJhon CucanchónÎncă nu există evaluări
- Fasciola HepaticaDocument3 paginiFasciola HepaticaAxoncomunicacion100% (1)
- 1.5 HanaDocument49 pagini1.5 HanaRosa Maria BarbozaÎncă nu există evaluări
- Carol LynneDocument96 paginiCarol Lynnediana100% (2)
- Dieta Restringida en PurinasDocument7 paginiDieta Restringida en Purinasmarta5358Încă nu există evaluări
- Alteraciones As Por AmalgamasDocument58 paginiAlteraciones As Por AmalgamasÁngel Soto RendonÎncă nu există evaluări
- De La Granja A La MesaDocument28 paginiDe La Granja A La MesaFrancis Figueroa LazaroÎncă nu există evaluări
- FISIOTERAPIA y Trastornos TemporomandibularesDocument125 paginiFISIOTERAPIA y Trastornos TemporomandibularesEvelyn CaizachanaÎncă nu există evaluări
- Conducta Del TigreDocument3 paginiConducta Del TigreelizabethÎncă nu există evaluări
- El Ocaso Del LoboDocument18 paginiEl Ocaso Del LoboErnesto GarcíaÎncă nu există evaluări
- CélulasDocument3 paginiCélulasYael Anahi CapitanÎncă nu există evaluări
- TERATOGENOSDocument21 paginiTERATOGENOSOskar Adrian Diaz VillanuevaÎncă nu există evaluări
- Pez CebraDocument6 paginiPez CebraGabrielCamarena100% (1)
- Capítulo 3 MalezasDocument15 paginiCapítulo 3 MalezasFrancklin CruzÎncă nu există evaluări
- El Libro de La Selva PDFDocument375 paginiEl Libro de La Selva PDFWILFREDO CRISPIN GUILLENÎncă nu există evaluări
- Anatomia I EsfenoidesDocument21 paginiAnatomia I EsfenoidesFernanda MadrigalÎncă nu există evaluări
- DRA MERCEDES PEñADocument17 paginiDRA MERCEDES PEñAJesus MartinezÎncă nu există evaluări
- Programa Islas Guañape 1Document2 paginiPrograma Islas Guañape 1Leedy Pacherres OsorioÎncă nu există evaluări
- Grado TerceroDocument67 paginiGrado Tercerosamu0225Încă nu există evaluări
- Multiples Lenguajes Sexto GradoDocument5 paginiMultiples Lenguajes Sexto GradoBerenice RamirezÎncă nu există evaluări
- Expo OrnitologiaDocument13 paginiExpo OrnitologiaEYVY MICHELL FLOREZ SALAMANCAÎncă nu există evaluări